Determinace pohlaví u krytosemenných rostlin
Pohlavní rozmnožování je nejrozšířenějším způsobem rozmnožování vyšších rostlin. kKvět s rozmnožovacími orgány má téměř neomezenou rozmanitost morfologických a fyziologických znaků, jejichž kombinace představuje různá přizpůsobení květů především k efektivnějšímu opylení a regulaci stupně cizosprášení. Jedním z takových přizpůsobení je i různé rozmístění pohlavních orgánů v květech, na rostlinách a ve skupinách rostlin. Při klasifikaci je třeba rozlišovat typy pohlaví v samostatných květech, na celých rostlinách a v rámci skupiny rostlin (obr. 6.1).
Typy pohlaví jednotlivých květů
- Oboupohlavný (obojaký, hermafroditní, hermaphroditus) – květ, který má jak tyčinky, tak i jeden nebo několik plodolistů
- Tyčinkový (samčí, masculum) – květ, který má pouze tyčinky a nemá plodolisty
- Pestíkový (samičí, feminus) – květ, který má pouze plodolisty a nemá tyčinky
Typy pohlaví jednotlivých rostlin
- Oboupohlavná (obojaká, hermafroditní, hermaphroditus) – rostlina, která má pouze oboupohlavné květy
- Jednodomá (monoecická, monoická, monoecious) – rostlina, která má jak tyčinkové, tak pestíkové květy
- Samčí (androecická, androecious) – rostlina, která má pouze tyčinkové květy
- Samičí (gynoecická, gynoecious) – rostlina, která má pouze pestíkové květy
- Andromonoecická (andromonoecious) – rostlina, která má jak oboupohlavné, tak tyčinkové květy (Veratrum album, Astrantia major)
- Gynomonoecická (gynomonoecious) – rostlina, která má jak oboupohlavné, tak pestíkové květy (Paraetaria officinalis)
- Trimonoecická (trimonoecious) – rostlina, která má tři typy květů: oboupohlavné, tyčinkové a pestíkové (Saponaria ocimoides)
- Agamonoecická (agamonoecious) – rostlina, která má vedle oboupohlavných květů ještě květy bezpohlavné se zakrnělými nebo abortovanými tyčinkami a semeníky (Viburnum opulus)
Typy pohlaví skupiny rostlin (populace, druhu, odrůdy)
- Oboupohlavná (obojaká, hermafroditní, hermaphroditus) – skupina, která má pouze oboupohlavné rostliny
- Jednodomá (monoecická, monoická, monoecious) – skupina, která má pouze jednodomé rostliny (Zea mays, Juglans regia, Corylus avelana)
- Dvoudomá (dioecická, dioická, dioecious) – skupina, která má samčí a samičí rostliny, (Bryonia dioica, Melandrium album, Silene alba, Humulus lupulus)
- Androdioecická (sub-androecická, androdioecious) – skupina, která má rostliny oboupohlavné a samčí (Dryas octopetala)
- Gynodioecická (sub-gynoecická, gynodioecious) – skupina, která má rostliny oboupohlavné a samičí (Origanum vulgare, Cirsium oleracea, Thymus serpyllum)
- Trioecická – skupina, která má rostliny oboupohlavné, samčí i samičí (Fraxinus excelsior, Asparagus officinalis)
Přehled uvedených typů a používané symboly uvádí obr. 6.1.
Výčet typů v uvedeném přehledu není vyčerpávající a existují ještě další možnosti či varianty, které by vyžadovaly zvláštní popis. Jako příklad lze uvést tři varianty jednodomé rostliny:
- tyčinkové a pestíkové květy jsou rozmístěny po celé rostlině (např. četné odrůdy okurek),
- tyčinkové a pestíkové květy se shlukují v samčí a samičí květenství (např. kukuřice) a
- v určitém stadiu růstu má rostlina květy pouze jednoho typu, jako např. u Arisaema japonica se zpočátku objevují pouze tyčinkové květy, později pouze pestíkové, takže se při zběžném pohledu jeví rostliny jako dvoudomé.
Podstatné je přesné vymezení pohlaví květů, rostlin a skupiny rostlin (např. musíme mluvit o tyčinkovém květu, samčí rostlině a dvoudomé populaci).
Převážná většina rostlin jsou rostliny oboupohlavné. Uvádí se, že mezi přibližně 120 tisíci druhy krytosemenných rostlin je 86 % druhů s rostlinami, které mají pohlavní orgány obou typů, 4 % druhů s rostlinami odděleného pohlaví a 10 % druhů ostatních. Je vidět, že jednopohlavných forem není v rostlinné říši mnoho. Toto rozdělení je významné z evolučního hlediska, neboť oboupohlavnost spolu s mechanizmy izolace různého typu dává více možností ve smyslu adaptace k měnícím se podmínkám prostředí než jednopohlavnost.
U kulturních rostlin se pohlavní dimorfizmus vyskytuje pouze u několika málo druhů; u mnoha dvoudomých kultur se objevují pravidelně nebo občas oboupohlavné nebo jednodomé rostliny v četnosti závislé na genetických faktorech či vnějších podmínkách. Dvoudomost se u rostlin hospodářsky významných vyskytuje hlavně u víceletých plodin. Jsou to např. Cannabis sativa, Simondsia californica, Humulus lupulus, Piper nigrum, Dioscorea sp., Spinacia oleracea, Asparagus officinalis, Poa arachnifera, Atriplex sp., Ilex aquifolium, Acer negundo, Fraxinus sp., Pistacia vera, Carica papaya, Vitis rotundifolia, Morus alba, M. nigra, Ficus carica a Phoenix dactylifera. U těchto rostlin dochází více nebo méně často k obligátnímu cizosprášení v závislosti na výskytu oboupohlavných nebo jednodomých rostlin v populacích a na stupni jejich inkompatibility.
U oboupohlavných rostlin můžeme pozorovat velkou rozmanitost ve způsobu rozmnožování. Takovéto rozdíly jsou podmíněny jak genotypovou variabilitou uvnitř druhů, tak vnějšími podmínkami, kterým je tato potenciálně cizosprašná populace vystavena. Podle způsobu rozmnožování se rostliny rozdělují na samosprašné (autogamní) a cizosprašné (alogamní).
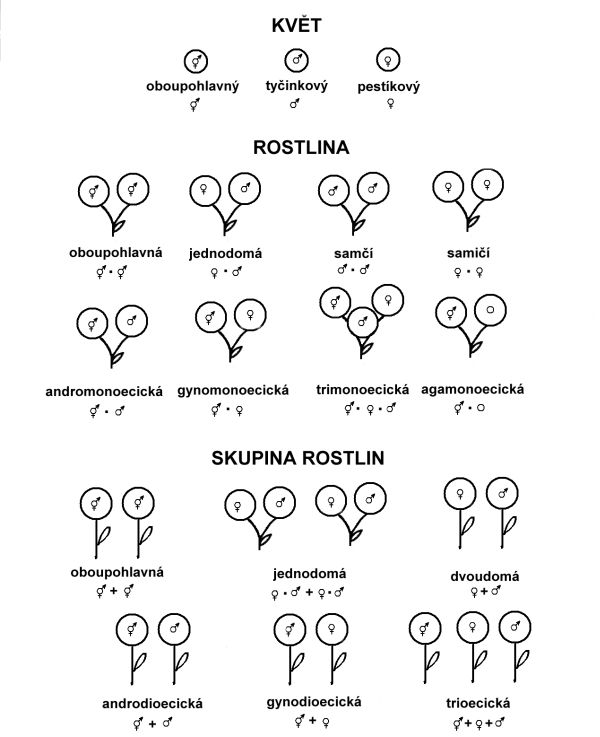
Obr. 6.1 Schematické znázornění a názvy jednotlivých typů pohlaví u květů, rostlin a skupin rostlin
(Zdroj: Řepková, Relichová, 2001)
Samosprašné jsou takové rostliny, u kterých dochází nejen k opylení, ale i oplození vlastním pylem a podobně u cizosprašných rostlin předpokládáme nejen opylení, ale i oplození cizím pylem. Proto je výstižnější hovořit o rostlinách se samooplozením a náhodným oplozením.
Determinace pohlaví
Genetické založení pohlaví u rostlin bylo experimentálně prokázáno brzy po znovuobjevení Mendelových principů dědičnosti. Jedním z těch, kteří se o to zasloužili, byl Carl Correns, který svoje závěry založil na výsledcích tří základních pokusů s křížením rostlin rodu Bryonia. V prvním případě křížil samičí rostliny B. dioica s jednodomými rostlinami B. alba a dostal 11 rostlin, které byly buď zcela samičí nebo samičí s několika brzy odumírajícími nebo nefunkčními tyčinkovými květy. Ve druhém pokusu křížil Correns samičí rostliny B. dioica se samčími rostlinami téhož druhu a získal 41 kvetoucích rostlin (21 samčích a 20 samičích). Ve třetím pokusu křížil rostliny B. alba se samičími květenstvími se samčími rostlinami B. dioica a získal 76 kvetoucích rostlin, polovinu samčích a polovinu samičích. Correns ještě tehdy neznal význam vnějších podmínek na projev znaku, proto některé výsledky nemohl spolehlivě vysvětlit. Jasně však poukázal na vztah mezi pohlavím a genotypem rostliny.
Objev pohlavních chromozomů u rostlin se datuje do roku 1923, kdy Blackburn, Kihara a Ono a nezávisle na nich Winge objevili heteromorfní bivalenty u Silene latifolia (Melandrium album), Rumex acetosa a Humulus sp. Poté byly pohlavní chromozomy objeveny u mnoha dalších dvoudomých rostlin. Přitom se však ukázalo, že ne všechny dvoudomé rostliny musí obsahovat tento heteromorfní bivalent.
Determinace pohlaví u krytosemenných rostlin je proces, který zajišťuje potlačení vývoje samčích nebo samičích orgánů v jednotlivých květech. V případě dvoudomých druhů dochází takto ke vzniku jedinců nesoucích pouze samčí nebo samičí květy. K potlačení vývoje orgánů opačného pohlaví dochází v různých fázích vývoje květu (obr. 6.2). Tento proces je ovlivněn geneticky; u některých druhů však může být ovlivněn také různými faktory vnějšího prostředí, jako je např. intenzita světla, délka dne, teplota a minerální výživa. Exogenní aplikace rostlinných hormonů také vede v některých případech k ovlivnění pohlaví květů. Žádná z těchto látek nemá shodný účinek u všech druhů s jednopohlavnými květy.
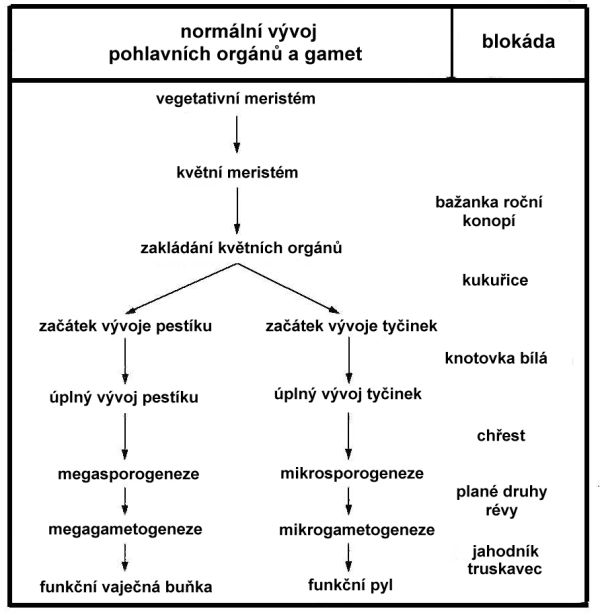
Obr. 6.2 Jednotlivé fáze potlačení vývoje orgánů určitého pohlaví u dvoudomých rostlin
(Zdroj: Janoušek, 1996)
Roku 1956 Hartmann formuloval tzv. „zákon potenciální oboupohlavnosti“, podle kterého nesou obě pohlaví veškerou genetickou informaci potřebnou k vytvoření pohlavních orgánů opačného pohlaví. Tento zákon odráží situaci u evolučně málo pokročilých typů determinace pohlaví, které jsou založeny na působení jednoho nebo několika genů řídících pohlaví. U druhů s pohlavními chromozomy se však setkáváme s tendencí k akumulaci genů, které jsou výhodné pouze pro heterogametické pohlaví, na chromozomu Y. Může proto docházet k výjimkám z tohoto zákona, neboť u homogametického pohlaví nemusí být potom přítomny všechny geny potřebné pro vznik orgánů opačného pohlaví. U pohlaví heterogametického však platí tento zákon bez výjimek.
Obecná teorie genetické determinace pohlaví vychází z předpokladu, že geny zapojené do exprese pohlaví je možné rozdělit do dvou skupin. Početná je první skupina genů, které jsou přímo zapojeny do morfogeneze pestíků nebo tyčinek (angl. sex differenciation genes). Druhou skupinu tvoří geny, které řídí expresi genů první skupiny (angl. sex determination genes).
Determinace pohlaví u jednodomých druhů
Jedním z možných modelů pro výzkum determinace pohlaví u rostlin je kukuřice, která je nejprozkoumanějším jednodomým druhem, ale podařilo se u ní získat i jednopohlavné linie. U kukuřice standardního fenotypu jsou samčí květy soustředěny v terminální latě (angl. tassel) a květy samičí se zase vyskytují pouze v postranních palicích (angl. ear), jejichž pestíky se vyznačují dlouhými bliznami (angl. silk). U kukuřice byl proveden pokus o vytvoření umělého systému dvoudomosti založeného na působení recesivních mutantních alel silkless1 (sk1) a tasselseed2 (ts2). Přítomnost alely sk1 v homozygotním stavu vede k potlačení vývoje pestíků samičích květů v postranních klasech a vzniká rostlina samčího fenotypu. Pokud je rostlina homozygotní pro alelu ts2, dochází u ní k přeměně všech samčích květů v latě v samičí, a proto opět vzniká jednopohlavná rostlina. K vytvoření stabilní dvoudomé populace mělo vést křížení rostlin genotypu sk1 sk1 ts2 ts2 (předpokládaný samičí fenotyp) se samčími rostlinami genotypu sk1 sk1 Ts2 ts2, jež by dalo vznik samčím a samičím rostlinám v poměru 1 : 1. Lokusy silkless1 a tasselseed2 však spolu interagují, a proto se rostliny genotypu sk1 sk1 ts2 ts2 vyznačovaly zcela neočekávaným fenotypem; v postranních palicích se u všech květů vyskytovaly pestíky a v koncovém květenství se kromě očekávaných samičích květů vyskytovaly také květy samčí a hermafroditní.
V současné době je již známo několik lokusů důležitých pro orgánově specifickou aborci, která je nezbytná pro vývoj jednopohlavných květů u kukuřice. Dva lokusy zapojené do suprese tvorby pestíků v terminálním květenství byly identifikovány pomocí recesivních mutací tasselseed (ts1 a ts2) a jeden lokus pomocí dominantní mutace Tasselseed5 (Ts5). U mutantů dochází k feminizaci tyčinkových květů a květy jsou oboupohlavné. Gen TS2 byl izolován prostřednictvím inzerční mutageneze; kóduje alkoholdehydrogenázu, která se podílí na buněčné smrti primordií pestíku. Genů, které se podílejí na řízení procesu aborce tyčinek je známo celkem sedm. Šest z nich jsou recesivní mutace: dwarf (d1, d2, d3, d5), anther ear (an1) a silkless (sk1). Jedna mutace je dominantní povahy – Dwarf8 (D8). Gen D8 je negativní regulátor kyseliny giberelové a u mutanta dochází k obnovení signálu, což má vliv na přerušení aborce tyčinek. V dalším výzkumu se ukázaly být velmi cenné rostliny recesivně homozygotní v lokusech dwarf (d1, d2, d3, d5), protože bylo zjištěno, že narušení funkce kteréhokoli z těchto lokusů vede k výraznému snížení obsahu giberelinů a maskulinizaci pestíkových květů, kdy je přerušena aborce tyčinek. Standardní alely genů se tedy podílejí na biosyntéze giberelinů a potlačení tvorby tyčinek.
Ve studiu genetické determinace pohlaví se využívá několik modelových druhů, které zastupují různé typy determinace pohlaví. K perspektivním modelovým objektům patří kukuřice setá (Zea mays), bažanka roční (Mercurialis annua) a knotovka bílá (Silene latifolia).
Na základě studia vzájemných interakcí různých lokusů byl navržen model, podle kterého jsou v procesu determinace pohlaví u kukuřice rozhodující interakce mezi lokusy TASSELSEED2, SILKLESS1 a hladinou giberelinů. Podle tohoto modelu standardní alela lokusu Sk1 blokuje působení produktu genu TASSELSEED2, a to vede k tvorbě pestíků v květech postranních klásků. V koncovém květenství je zase naopak působení genu SILKLESS1 blokováno genem TASSELSEED2, a proto zde dochází k aborci pestíků. Aborce tyčinek v samičím klasu u homozygotů TS2 TS2 SK1 SK1 je důsledek antagonistického působení pestíků na tyčinky prostřednictvím feminizujícího faktoru, jakým jsou gibereliny.
Typické zástupce jednodomých rostlin najdeme v čeledi Cucurbitaceae. Jeden z nejdůležitějších hospodářských druhů této čeledě je Cucumis sativus (dýně okurka). Snad u žádné jiné kultury nebyl prozkoumán projev pohlaví tak detailně, jako právě u okurky. Kromě toho je tento objekt modelovým systémem ke zkoumání morfogeneze vyšších rostlin. Proto také genetická kontrola jednodomosti může být vyložena na příkladu okurky.
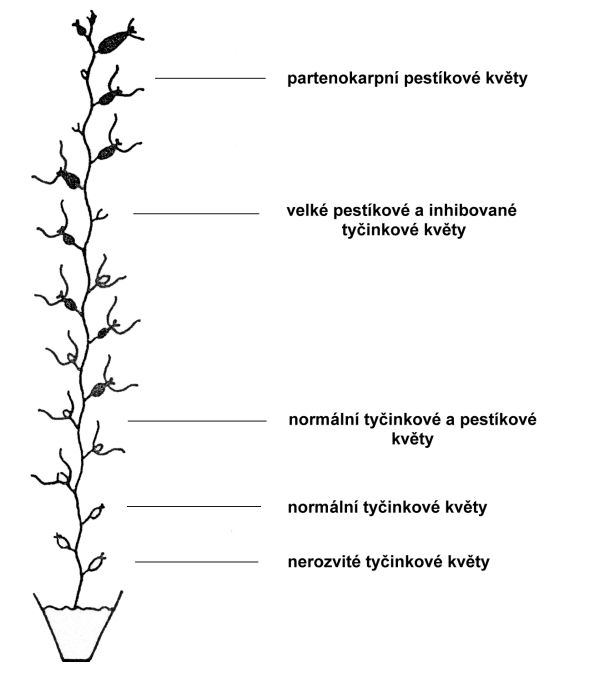
Rostliny okurky se vyznačují velkou různorodostí v projevu pohlaví (obr. 6.3). Květní poupata v raných fázích vývoje mají základy jak tyčinek, tak pestíků, avšak květy jsou většinou různopohlavné. Jednotlivé rostliny se vyznačují velkou variabilitou v rozmístění a počtu tyčinkových a pestíkových květů. U jednodomých rostlin okurky je prakticky ve všech úžlabích listů buď několik samčích, nebo jednotlivé (zřídka dva a více) samičí květy. Tendence k feminizaci je větší v akropetálním směru (podobně jako u Cucurbita pepo).
Některé odrůdy okurek mohou mít na rostlině kromě samčích květů i květy hermafroditní. Květy různého pohlaví se výrazně liší morfologicky (obr. 6.4).
Obr. 6.3 Schéma rozdělení květů různých typů na hlavním stonku Cucurbita pepo cv. Acorn
(Zdroj: Řepková, Relichová, 2001)
Determinace pohlaví u okurky se účastní tyto hlavní geny:
- M/m kontroluje spouštěcí mechanizmus v květním poupěti; v přítomnosti alely M, která je úplně dominantní, se tvoří pouze jednopohlavné květy;
- F/f kontroluje schopnost k vývinu pestíku (FF) a tvoří gradient pro samiččí pohlaví podél lodyhy; mezi alelami je vztah neúplné dominance;
- A/a gen je epistatický vůči genu F, zvyšuje samičí tendenci.
Obr. 6.4 Typy květů u Cucumis sativus
(Zdroj: Řepková, Relichová, 2001)
Kombinace těchto tří hlavních genů zodpovědných za determinaci pohlaví mohou dát následující fenotypy:
- M- FF populace monoecická, květy pestíkové
- M- ff populace monoecická, květy tyčinkové
- mm ff populace andromonoecická
Projev pohlaví u okurky může v některých případech podléhat i působení dalších genů.
Důležité je, že na projev pohlaví mají značný vliv i vnější podmínky, a to do té míry, že identické genotypy mohou dávat v různých podmínkách různé fenotypy a naopak různé genotypy mohou mít shodné fenotypy. Kromě klimatických podmínek (délka dne a teplota) mohou projev pohlaví ovlivňovat růstové regulátory a jiné chemické látky. Působením chemických látek lze změnit pohlaví libovolným směrem.
Systémy dvoudomosti založené na jednotlivých lokusech
Dvoudomost se vyskytuje v rostlinné říši asi dvakrát méně než jednodomost. Zcela dvoudomých je pouze asi 5 % čeledí a 75 % čeledí obsahuje některé dvoudomé druhy.
Pozornost byla dosud soustředěna převážně na druhy s pohlavními chromozomy, a proto jsou v současné době známy pouze dva druhy, u kterých je vznik samčích a samičích rostlin řízen jednotlivými lokusy: tykvice stříkavá (Ecballium elaterium) a bažanka roční (Mercurialis annua).
Typickým zástupcem této skupiny je druh tykvice stříkavá z čeledě tykvovitých (Cucurbitaceae), u které o expresi pohlaví rozhoduje přítomnost tří druhů alel (aD, a+, ad) v lokusu určujícím pohlaví. Alela aD je dominantní vůči alele a+ i alele ad, její přítomnost vede u rostlin k samčímu fenotypu. Pokud se uplatní alela a+, která je recesivní vzhledem k alele aD a dominantní vůči alele ad, vzniká rostlina jednodomá. Vznik samičích rostlin je podmíněn přítomností alely ad v homozygotním stavu.
Pohlaví řízené více lokusy
Morfogeneze tyčinek a pestíků je determinována větším počtem genů. U oboupohlavných rostlin jsou všechny geny potřebné k tvorbě obou typů květních orgánů přítomné a funkční. V procesu diferenciace (tj. při vývoji tyčinek nebo pestíků) jsou tyto geny různě exprimovány. Ztráta formy nebo funkce může být pak způsobena potlačením exprese příslušného genu nebo jeho nepřítomností.
U dvoudomých rostlin vzniknou při křížení mezi samičími a samčími rostlinami zase buď samičí, nebo samčí rostliny. Za předpokladu, že u obou pohlaví jsou přítomné jak geny zodpovědné za tvorbu tyčinek (A, B, C), tak geny pro vznik pestíků (D, E, F), pouze jejich aktivita je různá, odpovídá následující schéma:
Je vidět, že klíčovou roli hraje alelový pár M/m: mm podmiňuje expresi pouze těch genů, které jsou zodpovědné za tvorbu pestíků, Mm pouze za tvorbu tyčinek. Podle tohoto modelu může být diferenciace tyčinek a pestíků podmíněna neomezeným počtem genů, avšak determinace pohlaví je kontrolována pouze jedním lokusem působícím jako jakýsi spouštěcí mechanizmus (angl. trigger).
U bažanky roční je pohlaví determinováno třemi lokusy A/a, B1/b1 a B2/b2, které nevykazují genetickou vazbu. Pro vývoj samčího pohlaví je nutná přítomnost dominantní alely lokusu A/a a navíc alespoň v jednom z lokusů B. Dominantní alela lokusu B1/b1 má silnější maskulinizační efekt než alela B2. Zároveň se samčí a samičí rostliny vyznačují rozdílným obsahem rostlinných hormonů. Cytokininový hormon trans-zeatin je přítomen pouze u samičích rostlin, příbuzný cytokininový hormon trans-zeatin mononukleoid se vyskytuje pouze u samčích rostlin. Exprese pohlaví může být u jednoho druhu ovlivněna rostlinnými hormony v souladu s výše uvedenými rozdíly. Auxiny mají maskulinizující a cytokininy feminizující vliv.
Vícelokusovému modelu odpovídá experimentální navození dvoudomosti u kukuřice. U kukuřice bylo nalezeno mnoho mutantů, u kterých jsou potlačena samičí květenství (rostliny silkless) nebo změněna samčí květenství – laty tak, že místo nich se tvoří pestíkové květenství (tzv. tassel seed).
Alelový pár Ts2/ts2 můžeme v souhlase s předchozím modelem považovat za spouštěcí mechanizmus (trigger). Kromě genů Ts2/ts2 jsou ostatní geny determinující vývoj pohlavních orgánů pro obě pohlaví společné a homozygotní a je tedy lhostejné, kde jsou lokalizovány (rekombinace je neúčinná). V tomto modelu tvoří samčí rostliny vždy dva druhy gamet, zatímco samičí rostliny pouze jeden druh.
Umělá dvoudomost u kukuřice byla také navozena pomocí jiné kombinace genů. Gen ba (barren stalk) je recesivní, lokalizovaný na III. chromozomu a genotyp ba ba tvoří na rostlinách pouze laty. Podobně jako v předchozím případě budou rostliny ba ba ts2 ts2 samičí a ba ba Ts2 ts2 samčí. Podobně jako gen Ts2/ts2 může působit i jiný gen Ts3/ts3, v tomto případě však budou heterogametní rostliny samičí (ba ba Ts3 ts3). I když tyto systémy u Zea mays jsou velmi známé a často se uvádějí jako příklady umělé dvoudomosti, v některých případech se ukazuje tento systém složitější.
Jiný, složitější systém umělé dvoudomosti byl nalezen u Rubus idaeus. Předpokládá supresorové geny a vazbu mezi geny. Při determinaci pohlaví u normálně hermafroditního druhu R. idaeus působí dva geny: jeden potlačuje diferenciaci tyčinek (M/m) a druhý diferenciaci pestíků (F/f). Přitom samičí supresor je epistatický nad samčím supresorem. Tzn., že genotypy FF MM, Ff Mm a FF Mm budou samčí rostliny a genotyp ff mm samičí. Mezi oběma geny existuje navíc silná vazba. V tomto případě křížení ♀ fm fm x ♂ FM fm dá pouze rodičovské typy, což odpovídá systému dvoudomosti u R. idaeus.
Popsaný model se může zdát složitý, představuje však obecný princip pohlavního dimorfizmu u rostlin. Jiným způsobem zábrany crossing-overu mezi dvěma protichůdnými supresorovými geny je mechanická bariéra na chromozomové úrovni, tj. chromozomová aberace.
Systémy determinace pohlaví u rostlin s pohlavními chromozomy
Pohlavní dimorfizmus založený na supresorových genech se bude nejefektivněji projevovat při vazbě těchto genů. To vedlo ke vzniku pohlavních heteromorfních chromozomů, které se zřejmě vyvíjely v pozdějších stadiích evoluce a pouze u těch druhů, u kterých existovala dvoudomost. U mnohých dvoudomých druhů je heteromorfizmus pohlavních chromozomů morfologicky vyjádřen slabě a může být cytologicky odhalen pouze v určitých fázích meiózy. Navíc se stupeň tohoto dimorfizmu může i u velmi blízkých dvoudomých druhů značně lišit, což často vedlo k nejednotným experimentálním závěrům. V současné době byl dimorfizmus pohlavních chromozomů bezpečně prokázán u 13 druhů rostlin ze tří rodů (někteří autoři uvádějí 70 druhů z 24 rodů).
Nejjednodušším případem dimorfizmu chromozomů je systém X/Y. Tyto pohlavní chromozomy musí obsahovat alespoň dva odlišné segmenty: jeden homologní, který umožňuje synapsi v meióze a druhý heterologní. Složitější systémy zahrnují několik chromozomů X a Y. Zvláštním případem je systém X/0, jehož existence byla zjištěna pouze u tří druhů rostlin patřících k rodu Dioscorea, avšak některými autory nebyla potvrzena.
V tab. 6.1 jsou uvedeny druhy rostlin s heterologními pohlavními chromozomy a je z ní zřejmé, že v převážné většině případů jsou heteromorfní chromozomy u samčího pohlaví. Pouze u Fragaria elatior je heteromorfní samičí pohlaví. V rodu Fragaria se vyskytuje dvoudomost pouze na polyploidní úrovni (F. orientalis – 4x, F. elatior – 6x, F. virginiana, F. grandiflora – 8x).
| Druh | Pohlavní chromozomy | |
| ♀ | ♂ | |
| Cannabis sativa | XX | XY |
| Fragaria elatior | XY | XX |
| Humulus lupulus | XX | XY |
| H. lupulus var. cordifolius | X1X1X2X2 | X1Y1X2Y2 |
| H. japonicus | XX | XY1Y2 |
| Rumex angiocarpus | XX | XY |
| R. acetosa | XX | XY1Y2 |
| R. paucifolius | XX | XY |
| Silene latifolia | XX | XY |
| Melandrium rubrum | XX | XY |
| Populus sp. | XX | XY |
| Urtica dioica | XX | XY |
| Spinacia oleracea | XX | XY nebo YY |
U rostlin s pohlavními chromozomy jsou zpravidla uváděny základní typy determinace pohlaví, které jsou reprezentovány dvěma modelovými objekty: knotovkou bílou (Silene latifolia) a šťovíkem kyselým (Rumex acetosa). Nejvíce pozornosti bylo dosud věnováno druhům, které se vyznačují přítomností heteromorfních pohlavních chromozomů (tab. 6.1).
Typ s aktivní úlohou chromozomu Y – Silene latifolia
U S. latifolia uskutečnil už Correns ve 20. letech technicky velmi jednoduché pokusy ke zjištění, které pohlaví je heterogametické. Tyto pokusy byly založeny na předpokladu, že heterogametické pohlaví produkuje dva druhy pylu, u nichž existuje konkurence. Ta se projeví při opylení dostatečným množstvím pylu tak, že v potomstvu vzniká více samičích rostlin než samčích. Naopak při opylení malým množstvím pylu se konkurence neuplatní a poměr pohlaví je zhruba 1:1. Podobné jednoduché pokusy byly založeny na rozdílném přežívání pylových zrn. Knotovka bílá má systém determinace pohlaví založený na aktivním působení chromozomu Y (samci se vyznačují konstitucí AAXY a samice AAXX), který obsahuje geny důležité pro tvorbu tyčinek a gen nebo genový komplex suprimující expresi samičího pohlaví. Chromozomy X a Y se u knotovky bílé navíc významně liší a exprese pohlaví nepodléhá vlivům prostředí. Kromě knotovky bílé a její příbuzné knotovky červené se tímto typem určení pohlaví vyznačují i některé jiné druhy z různých čeledí: chřest (Asparagus officinalis), plané druhy révy (Coccinia indica), šťovík menší (Rumex acetosella) a pravděpodobně i mnoho dalších dosud neprozkoumaných druhů s heteromorfními, případně i homomorfními pohlavními chromozomy. Chřest se vyznačuje homomorfními pohlavními chromozomy. Také funkční odlišnost mezi chromozomem X a Y není u tohoto druhu tak výrazná jako u dvoudomých druhů knotovek a je tedy možno získat i tzv. supersamce, kteří mají genetickou konstituci AAYY. Zatímco samičí rostliny nesou pouze rudimenty tyčinek, nejsou u samčích rostlin pestíky zcela zakrnělé a vykazují značnou variabilitu. Také tento fakt svědčí o menší evoluční pokročilosti systému determinace pohlaví u tohoto druhu a pravděpodobném vzniku dvoudomosti prostřednictvím gynodioecie. Dosavadní studie se zde zaměřovaly spíše na hledání pohlavně specifických rozdílů na úrovni proteinů. Z obecně genetického hlediska je zjevná podobnost determinace pohlaví u knotovky a typu savčího. Oba tyto systémy jsou založeny na aktivní úloze chromozomu Y, který nese geny potlačující expresi samičího pohlaví. Vlastní biochemické mechanizmy, které realizují expresi pohlaví, jsou však zásadně odlišné i vlivem celkově odlišného metabolizmu rostlin a savců.
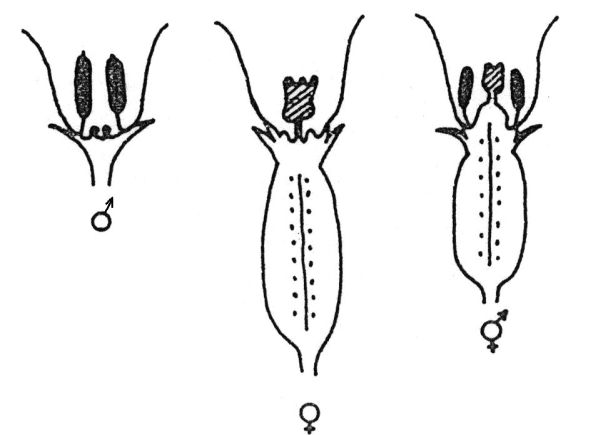
Studium rostlin s aberantními chromozomy, které prováděl Westergaard, ukázalo na různý význam jednotlivých segmentů chromozomu Y (obr. 6.5): segment I potlačuje vývoj samičích orgánů, avšak nepřekáží vývoji samčího pohlaví (rostliny XY1 mají květy se ♀ i ♂ orgány). Segmenty III a IV nepotlačují vývoj samičích orgánů, mají však vliv na konec etapy diferenciace prašníků (rostliny XY3 jsou samčí a pylově sterilní). Centrální segment II obsahuje geny, které kontrolují počáteční etapy tvorby prašníku. Chromozom Y je částečně homologní s chromozomem X (segment IV). Zbývající nehomologní část chromozomu X (segment V) obsahuje determinanty samičího pohlaví. Y1, Y2 a Y3 jsou skupiny rostlin z určité populace, které nesou delece různého rozsahu a polohy. Na základě hermafroditního fenotypu rostlin s chromozomem Y1 a Y2 bylo zjištěno, že diferenciální rameno chromozomu Y nese samičí supresor. Jelikož rostliny, které diferenciální rameno zcela postrádaly (typ Y2), tvořily normální prašníky, bylo možné konstatovat, že geny nezbytné pro tvorbu tyčinek (tzv. male promoter) leží v rameni částečně homologním s chromozomem X. V distální části homologního ramene chromozomu Y Westergaard dokázal přítomnost genů důležitých pro samčí fertilitu na základě fenotypu rostliny typu Y3.
U S. latifolia se sleduje možný vliv metylace DNA při determinaci pohlaví, protože byla pozorována hypermetylace jednoho nebo dvou chromozomů X. Nabízí se domněnka, že u rostlin by mohl existovat mechanizmus zajišťující kompenzaci funkčních alel uložených v pohlavních chromozomech X genetickou inaktivací jednoho nebo dvou chromozomů X u samičích rostlin a to metylací. Podobně je tomu u homogametického pohlaví savčího typu (tzv. lyonizace).
Podobně jako u S. latifolia je určeno pohlaví u R. acetosella, který však obsahuje homomorfní pohlavní chromozomy, označované jako SF a Sm. Chromozom Sm hraje rozhodující úlohu v determinaci pohlaví tím, že inhibuje samičí potenci lokalizovanou v chromozomu SF. Předpokládá se, že celkově je pohlaví kontrolováno několika málo geny s velkým účinkem a několika geny s malým účinkem lokalizovanými na uvedených chromozomech SF a Sm. Autozomy se nepodílejí na determinaci pohlaví.
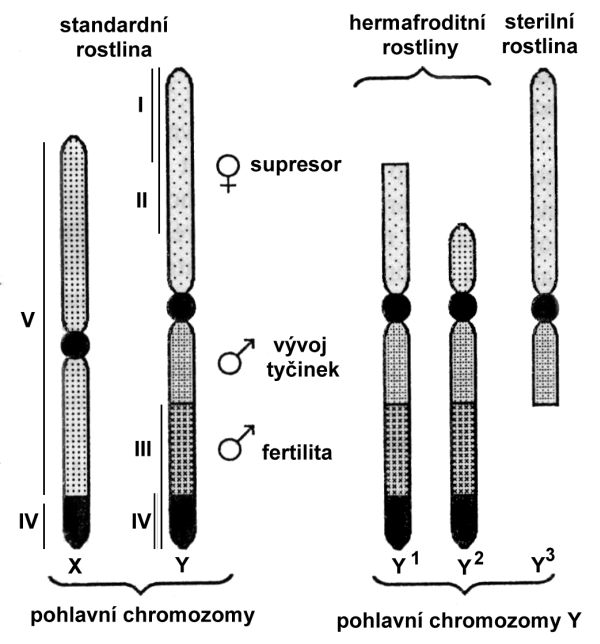
Obr. 6.5 Schematické znázornění normálních chromozomů X a Y u Silene latifolia a experimentálně získaných aberantních chromozomů Y1, Y 2 a Y3.
(Zdroj: Janoušek, 1996)
Typ s omezenou rolí chromozomu Y – Rumex acetosa
U druhů příbuzných šťovíku kyselému (Rumex acetosa) rozhoduje o expresi pohlaví poměr chromozomů X a sad autozomů. Tento pohlavní index je u samičích rostlin 1,0 (2A + XX) a u samčích rostlin 0,5 (2A + XY1Y2). V pokusech s polyploidy bylo zjištěno, že rostliny s poměrem 0,5 a menším, např. 3A + XY1Y2 byly samčí, zatímco rostliny s poměrem 1,0 a vyšším, např. 3A + XXXX byly samičí. Rostliny s poměrem od 0,5 do 1,0 zaujímají střední postavení (oboupohlavné rostliny s karyotypy 3A + XXY a 3A + XXYY mají pohlavní index 0,67).
Vývoj tyčinek není závislý na přítomnosti chromozomu Y. Exprese pohlaví je velmi stabilní. Ztráta role chromozomu Y se projevuje jeho značným polymorfizmem v populacích rostlin. U samotného šťovíku kyselého jsou dokonce přítomny v buňkách samčích rostlin současně s chromozomem X také dva chromozomy Y, které vykazují opět značnou variabilitu. Chmel japonský (Humulus japonicus) se podobně jako šťovík kyselý vyznačuje přítomností dvou chromozomů Y v buňkách. Systém determinace pohlaví u typu šťovík je z obecně genetického hlediska podobný typu Drosophila, neboť ani zde nerozhoduje o expresi daného pohlaví přítomnost či nepřítomnost chromozomu Y, ale poměr počtu chromozomů X a sad autozomů.
Typ s heterogametickým samičím pohlavím
Heterogametnost samičích rostlin byla dosud popsána pouze u planých polyploidních druhů jahodníků a u silenky ušnice (Silene otites). U silenky ušnice to bylo prokázáno křížením uměle získaných polyploidních samčích a samičích rostlin s diploidními rostlinami. Pokud byla samičí rostlina tetraploidní, tedy konstituce AAAAXXYY, vyskytovaly se v potomstvu takřka výhradně rostliny samičí, neboť gamety XX a YY se tvoří pouze výjimečně a převládají gamety XY. Tento typ připomíná systém determinace pohlaví u ptáků, neboť i zde je heterogametním pohlavím pohlaví samičí.
On-line prezentace
Ke kapitole Determinace pohlaví u krytosemenných rostlin.
ÚEB Biol, Přírodovědecká fakulta, Masarykova univerzita |
Návrat na úvodní stránku webu, přístupnost |
| Servisní středisko pro e-learning na MU
| Fakulta informatiky Masarykovy univerzity, 2013
Centrum interaktivních a multimediálních studijních opor pro inovaci výuky a efektivní učení | CZ.1.07/2.2.00/28.0041
