Přírodovědecká fakulta Masarykovy univerzity
Ing. Martin Krsek, CSc.
Význam půdních mikroorganismů
Mikroorganismy hrají nezastupitelnou roli v životě lidské společnosti. Jsou využívány v potravinářství, humánní i veterinární medicíně i v nejrůznějších průmyslových aplikacích a také většina moderních biotechnologií je založena na mikrobiologických základech. Kromě této pozitivní role způsobují mikrobi lidstvu i mnoho komplikací, ať už ve formě patogenů živočichů i rostlin, či původců nejrůznějších ztrát (kažení potravin, koroze materiálů a podobně). V první řadě jsou ale mikrobi nepostradatelnou součástí všech ekosystémů. Jsou zodpovědní za koloběh prvků v přírodě a podílejí se na mnoha významných procesech v terestriálních i akvatických ekosystémech. Jsou zdroji živin v základně všech potravinových řetězců. Rozsáhlé degradační schopnosti mikrobů jsou ve stále větší míře využívány při bioremediacích pro odstraňování nejrůznějších polutantů. I pouhá biomasa mikroorganismů na Zemi je – i přes jejich mikroskopické rozměry – úctyhodná. Podle některých odhadů tvoří víc jak třetinu celkové biomasy na Zemi (Whitman et al., 1998). Značná část této biomasy se nachází v terestriálních ekosystémech, především v půdě (Tab. 1-1), která představuje základní prostředí zabezpečující produkci naprosté většiny suchozemské organické hmoty a nepřímo také akvatické organické hmoty.
Půda představuje heterogenní médium sestávající z pevné, tekuté a plynné fáze. Na tvorbě půdy se podílí pět vzájemně reagujících faktorů: klíma, topografie, matečný materiál (hornina), čas a v neposlední řadě půdní organismy, kde klíčovou roli plní půdní mikroorganismy. Rozpad matečné horniny na jemné částice doprovázený uvolňováním živin zahajuje tvorbu půdy. Vzhledem k nedostatku využitelného C a N v raných stádiích tvorby půdy připadá klíčová role mikrobům schopným vázat tyto elementy, tedy fotosyntetizujícím a dusík-poutajícím mikroorganismům, ke kterým se postupně přidávají další skupiny mikrobů, jejichž funkcí je využití a především degradace vytvořené organické hmoty a návrat všech prvků do přirozeného koloběhu látek.
Odhady, kolik je na Zemi půdy, se liší. Vyjděme z předpokladu, že z celkového povrchu Země zhruba 25 % připadá na pevninu a jen polovina z této výměry je vhodná pro nějakou rostlinnou produkci (zbytek je tvořen pouštěmi, polárními oblastmi, nebo prostředím, kde je příliš teplo nebo chladno či jinak nehostinně). Zhruba 40 % z těchto 12,5 % je příliš limitováno terénními podmínkami, úrodností oblasti, či příliš vysokými srážkami, aby podporovaly jakoukoliv produkci potravin. To znamená, že k produkci potravin zbývá zhruba pouhých 7,5 % plochy zeměkoule, o které ještě soutěží s ostatními potřebami lidské společnosti, jako je budování měst, komunikací, průmyslových podniků a podobně. I přesto – nebo právě proto – představuje těch několik procent povrchu Země významné prostředí, jehož studiu se věnuje mnoho oborů lidské činnosti a to nejen z důvodů produkce potravin a dalších surovin. Půda také představuje prostředí filtrující vodu (včetně odpadních vod), produkuje i absorbuje významná množství plynů, je materiálem pro mnohá další odvětví lidské činnosti (produkce stavebnin, využití v lékařství, umělecké činnosti), je domovem pro mnoho živočichů a dalších organismů a v neposlední řadě místem dekompozice většiny organických i anorganických „odpadních“ produktů. Ve většině zmíněných procesů hrají klíčovou roli mikroorganismy přítomné v půdě. Jejich hlavní funkcí je již zmíněná dekompozice organické hmoty (včetně těl mikrobů). Během tohoto procesu dochází zároveň k tvorbě nových mikrobiálních buněk i vytváření nových organických látek (včetně humusu). Přesto však představa o půdě kypící mikrobiálním životem je pro většinu půdního prostředí (s výjimkou rhizosferní půdy) s největší pravděpodobností mylná. Většina mikroorganismů je v půdě v klidovém stádiu čekající na svou příležitost, která nastane s přísunem nové organické hmoty.
| Organisms | Number | Biomass | ||
| per mm2 | per gramm | kg/ha | g/mm2 | |
| Microflora | ||||
| Bacteria | 1013–1014 | 108–109 | 400–5000 | 40–500 |
| Actinomyctetes | 1012–1013 | 107–108 | 400–5000 | 40–500 |
| Fungi | 1010–1011 | 105–106 | 1000–20 000 | 100–2000 |
| Algae | 109–1010 | 104–105 | 10–500 | 1–50 |
| Fauna | ||||
| Protoza | 109–1010 | 104–105 | 20–200 | 2–20 |
| Nematodes | 106–107 | 10–102 | 10–150 | 1–15 |
| Mites | 103–106 | 1–10 | 5–150 | 0,5–1,5 |
| Collembola | 103–106 | 1–10 | 5–150 | 0,5–1,5 |
| Earthworms | 10–103 | – | 100–1700 | 10–170 |
| Other fauna | 102–104 | – | 10–100 | 1–10 |
(Převzato z: Scow, 2000)
Stručná charakteristika skupin půdních mikroorganismů
Půdní mikroorganismy zahrnují širokou řadu funkčních a taxonomických skupin a najdeme zde zástupce všech tří fylogenetických domén Bacteria, Archaea a Eukarya (Tab. 1). Nejvýznamnější skupinu co do jejich počtu i biomasy představují bakterie a houby, přesto ale i ostatní skupiny půdních (mikro)organismů nelze přehlížet. Plní v půdním prostředí své nezastupitelné funkce a společně se podílí na správném fungování půdního prostředí, což nás téměř opravňuje odhlédnout od jednotlivých půdních organismů a uvažovat o půdě jako o velkém mnohobuněčném organismu.
Viry
Nejmenšími „organizmy“ v půdě jsou viry. Jedná se o obligátní parazity všech organismů nacházejících se v půdě. Tato skupina na pomezí živé a neživé hmoty nemá svůj vlastní metabolismus (a proto často viry ani nejsou zařazovány mezi organismy) a naše znalosti o jejím zastoupení a funkci v půdě jsou více než kusé. Až do nedávné doby se studium virů v půdě omezovalo na bakteriofágy významných skupin půdních bakterií, například fixátorů vzdušného dusíku a podobně. Zvýšený zájem o půdní viry byl podnícen výsledky studií virů ve vodním, především mořském prostředí, které odhalily skutečnost, že množství virů zde převyšuje množství prokaryotických mikroorganismů se všemi z toho vyplývajícími důsledky (lytické působení virů, transdukce a podobně). Například Ashelford et al. (2003) uvádí hustotu populace virů v akvatickém prostředí 2,5×108 ml−1 převyšující tak 10× hustotu populace jejich bakteriálních hostitelů. Průměrný počet fágů z půdy získaný transmisní elektronovou mikroskopií stanovili Ashelford et al. (2003) na 1,5×107 g−1. Weinbauer a Rassoulzadegan (2004) shrnují poznatky a úvahy o úloze virů v akvatických systémech a zdůrazňují jejich význam v následujících oblastech: vliv na genetickou diverzitu mikrobiálních komunit, především jejich možnou roli v regulaci četností bakteriálních populací (teorie „killing the winner“, podle které by viry svým lytickým působením zabraňovaly „přemnožení“ kompetitivních druhů) a přenos genů mezi druhy. Podle těchto autorů geny a aktivita virů vytváří genetickou variabilitu prokaryot a je hnací silou ekologického fungování těchto komunit a jejich evolučních změn. Situace v půdním prostředí má svá specifika (mikroskopická struktura půdy s množstvím povrchů schopných poutat mnohé „složky“ půdního prostředí), přesto však funkce a význam virů v tomto prostředí budou velmi podobné prostředí akvatickému.
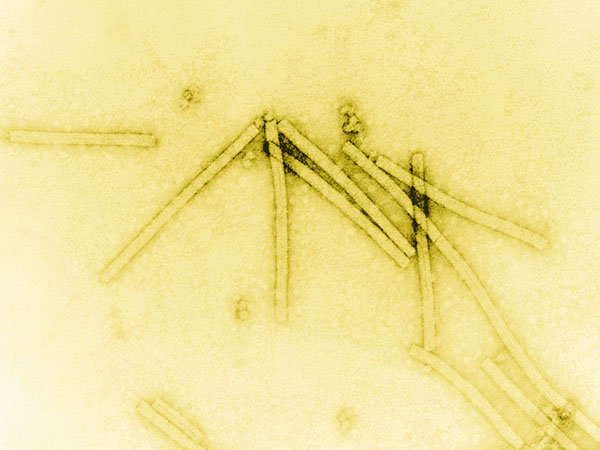
Obr. 1-1 Virus tabákové mozaiky
(zdroj: Wikimedia commons)
Nejzávažnější překážkou studia virů je nemožnost jejich kultivace bez jejich hostitele. Přidáme-li k tomu i známý a všeobecně uznávaný fakt, že například ze všech bakterií přítomných v životním prostředí je méně než 1 % kultivovatelných (Torsvik et al., 1990a), je jasné, že studium bakteriofágů v půdním prostředí je odkázané především na molekulárně biologické metody (a podobně tomu bude i u ostatních mikroorganismů v půdě – archeí i mikromycet). I zde však narážíme na další komplikace – malá velikost studovaných částic a silné vazby na pevnou složku půdy znesnadňující jejich extrakci z tohoto prostředí a především pak absence jakéhokoliv molekulárního markeru (obdoba genů 16S rDNA u prokaryotických organismů) umožňujícího detekci virů a studium jejich diverzity.
Významným přínosem k metodologii studia virů v půdním prostředí byla práce Williamsona et al. (2003), ve které autoři srovnávali různé pufry k extrakci fágů z půdy a v návaznosti na to techniky použité ke kvantifikaci fágů (počítání plak, epifluorescenční mikroskopii a transmisní elektronovou mikroskopii). Publikované údaje o četnosti virů v půdě se pohybují v závislosti na použité technice v rozmezí od 107 do 109 a tedy i v tomto prostředí jejich četnost pravděpodobně přesahuje (zhruba 10x) četnost jejich hostitelů, bakterií. První údaje získané studiem metagenomových knihoven naznačují, že většina genů dvouřetězcových DNA virů je neznámá. Pro srovnání zhruba 85 % mikrobiálních metagenomových sekvencí má významnou homologii s geny v GenBank, ale v případě fágových sekvencí je to pouze 35 % (Srinivasiah et al., 2008). Kromě volných virů nesmíme také zapomenout na temperované fágy. Výsledky různých studií naznačují, že zhruba 30 % kultivovatelných půdních bakterií je lyzogenních a podobná čísla zřejmě platí i pro nekultivovatelnou složku půdní mikroflóry (Williamson et al., 2007, 2008).
Archaea
Podobně informace o roli zástupců Archaea v půdě, s výjimkou zaplavených půd a jiných extrémních habitatů, jsou značně omezené. Až do nedávné doby se předpokládalo, že Archaea se nacházejí především v extrémních prostředích, nicméně ke konci 90. let minulého století se začaly objevovat studie potvrzující přítomnost zástupců domény Archaea v půdě i rhizosféře rostlin (Bintrim et al., 1997; Buckley et al., 1998; Simon et al., 2005; Nicol et al., 2003). Jedná se především o ne-exterémofilní zástupce skupiny Crenarcheota. Podíl archeí na celkovém počtu prokaryotických organismů v půdě byl pomocí analýzy 16S rRNA určen na 1,42 % ± 0,42 % (Buckley et al., 1998). Úloha archeí v půdním ekosystému není ještě jednoznačně určená, ale například studie Leiningera et al. (2006) odhalila významný podíl archeí na procesech nitrifikace. Studiem genů kódujících podjednotku klíčového enzymu amonia monooxygenaze (amoA) byli tito autoři schopni stanovit, že počet kopií genu amoA archeálního původu je 3000× vyšší než počet amoA bakteriálního původu.
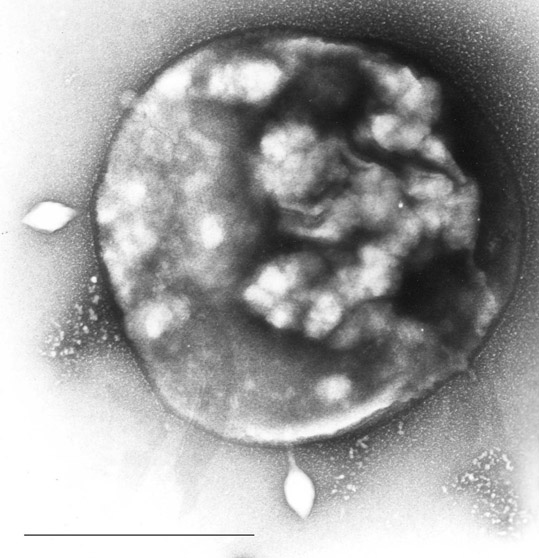
Obr. 1-2 Archaea rodu Sulfolobus
(zdroj: Wikimedia commons)
Bakterie
Nejvíce studovaným objektem půdní mikroflóry jsou bezesporu bakterie. V 1 g půdy se v závislosti na její úrodnosti i způsobu stanovení nachází 108 – 1010 bakteriálních buněk. Odhady počtů bakteriálních druhů nacházejících se v půdě se velmi liší. Encyclopedia of Microbiology (Scow, 2000) uvádí 13 tisíc bakteriálních druhů, Reid a Wong (2005) mluví o 60 tisících různých bakteriálních druhů přítomných v půdě. Podle výsledků renaturační kinetiky DNA bakteriální frakce extrahované z půdy může jeden gram půdy obsahovat 4000 zcela rozdílných geonomů standardních půdních bakterií (Torsvik et al., 1990a). S množstvím bakteriálních druhů zastoupených v půdě koresponduje i jejich metabolická variabilita nebo funkce, které v půdě plní. Základní funkcí je bezesporu degradace organických substrátů vstupujících do půdního prostředí, kde bakterie hrají – především v počátečních fázích rozkladu charakterizovaných dostatečnou vlhkostí substrátu – vedoucí roli (v pozdějších fázích rozkladu mohou dominovat houby). Významné místo zaujímají také symbiotičtí i volně žijící fixátoři vzdušného dusíku schopní doplnit tento základní makrobiogenní element v půdním prostředí, kde jeho nedostatek je často limitujícím faktorem všech fyziologických procesů v půdě, degradace nevyjímaje a také ostatní bakterie koloběhu dusíku (nitrifikační a denitrifikační). Na tomto místě by bylo možné zmínit i mnoho dalších skupin bakterií, jejichž role je pro správnou funkci půdního prostředí nezastupitelná, jako například aktinobakterie, síru oxidující bakterie a podobně.
Přehled nejčastějších zástupců půdních bakterií uvádí například všeobecně uznávaná Soil Microbiology and Biochemistry autorů Paula a Clarka (1989). Podle této publikace jsou numericky převládajícími půdními bakteriemi zástupci rodu Arthrobacter, který by podle některých odhadů měl představovat až 40 % bakterií detekovaných kultivačními metodami. Jedná se o pleomorfní grampozitivní až gramvariabilní pomalu rostoucí buňky, někdy pohyblivé. V množství mezi 5 až 20 % jsou v půdě často detekovány další tři rody: Streptomyces, Pseudomonas a Bacillus.
Rod Streptomyces představuje až 90 % zástupců řádu Actinomycetales. Jedná se o myceliární grampozitivní organismus množící se pomocí vzdušných spor nebo rozpadem mycelia. Najdeme ho na sušších stanovištích s neutrální až alkalickou reakcí. Tento rod (a i celý zmíněný řád) je znám produkcí četných antibiotik i dalších komerčně zajímavých látek (sekundárních metabolitů) a v neposlední řadě také geosminu zodpovědného za typickou vůni půdy.
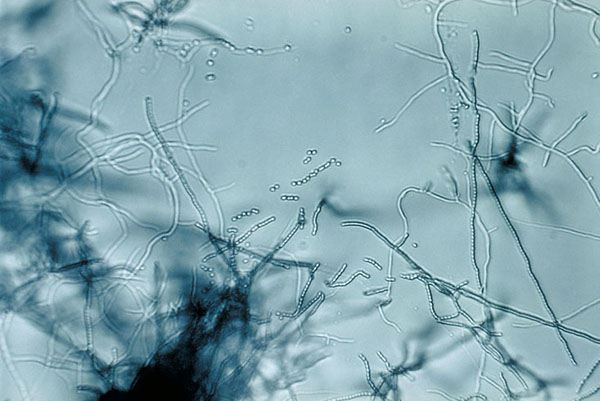
Obr. 1-3 Bakterie rodu Streptomyces
(zdroj: Wikimedia commons)
Rod Pseudomonas patří ke gramnegativním tyčinkovitým bakteriím, je opatřen polárním bičíkem a většina zástupců je aerobní (s výjimkou denitrifikačních bakterií využívajících nitráty jako konečný akceptor elektronů). Mnoho představitelů tohoto rodu žije v těsné asociaci s rostlinami, některé z nich mohou způsobit i choroby rostlin. Častá je u tohoto rodu produkce fluorescentních pigmentů. Siderofory produkované zástupci tohoto rodu mají silnou afinitu k Fe3+ a díky tomu mohou být použity k biologické kontrole půdních rostlinných patogenů.
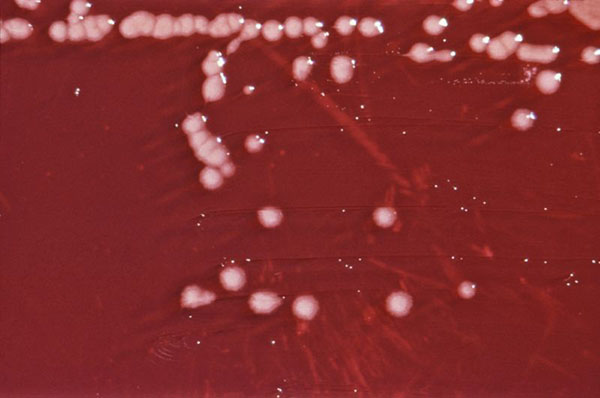
Obr. 1-4 Bakterie rodu Pseudomonas
(zdroj: Wikimedia commons)
Představitelé rodu Bacillus jsou grampozitivní sporulující většinou pohyblivé aerobní tyčinky. B. polymyxa je někdy uváděn jako fixátor atmosférického dusíku. Rod dále zahrnuje druhy používané pro biologickou ochranu rostlin (B. thuringensis), máčení lnu (B. macerans), ale i živočišné patogeny, jako B. anthracis.
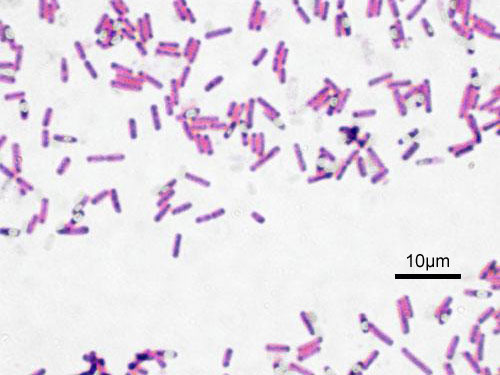
Obr. 1-5 Bakterie rodu Bacillus
(zdroj: Wikimedia commons)
Další častou půdní bakterií je rod Clostridium, opět sporulující avšak ve většině případů striktně anaerobní grampozitivní tyčinkovité bakterie. Také zde se vyskytují fixátoři dusíku, ale také druhy produkující silné toxiny (botulotoxin a tetanotoxin) a druhy patogenní pro živočichy včetně lidí. Jiné druhy mohou být použity pro produkci alkoholů a rozpouštědel.
Jak již bylo zmíněno výše, významnou skupinu půdních bakterií (ne tak svým početním zastoupením, jako svou funkcí v půdních mikrobiálních společenstvech) tvoří bakterie koloběhu esenciálního makrobiogenního prvku – dusíku. Patří sem v prvé řadě bakterie schopné poutat vzdušný dusík, tedy jednak volně žijící fixátoři, představovaní například rody Azotobacter, Azomonas, Beijerinckia, Derxia, Azospirillum a především pak symbiotičtí fixátoři rodů Rhizobium, Bradyrhizobium (některé kmeny tohoto rodu jsou schopné fixovat dusík i mimo rostliny) a Sinorhizobium. Na pomezí mezi volně žijícími a symbiotickými fixátory dusíku se nachází kmen Cyanobacteria, grampozitivní fototrofní bakterie, které se uplatňují zejména v prvotních stádiích tvorby půdy (najdeme je však i na povrchu již vytvořených půd), kdy jako primární kolonizátoři matečného materiálu uplatní svou schopnost fotosyntézy i fixace atmosférického dusíku ať už samostatně, nebo v symbióze s houbami v lišejnících. Z ostatních bakterií koloběhu dusíku je třeba zmínit nitrifikační bakterie oxidující ionty NH4+ za aerobních podmínek přes NO2− na NO3− (a tím převádějící relativně imobilní formu dusíku NH4+ na velmi pohyblivou a tedy lehce vymyvatelnou formu NO3-) reprezentované například rody Nitrosomonas, Nitrosolobus, Nitrospira, Nitrosovibrio a Nitrobacter a bakterie denitrifikační, které naopak za anaerobních podmínek mohou využívat oxidy dusíku (NO3− , NO2−, NO a N2O) jako akceptory elektronů a převádět je zpět až na vzdušný dusík N2. Schopnost denitrifikace má celá řada bakterií, například i zástupci výše zmíněných rodů Pseudomonas, Bradyrhizobium a další. Procesy koloběhu dusíku jsou významné nejen z hlediska „hospodaření“ s tímto esenciálním makrobiogenním prvkem, ale například uvolnění N2O během procesů přeměny dusíku představuje významný zdroj skleníkových plynů.
Je však třeba mít na paměti, že většina zvláště dříve publikovaných studií půdních mikroorganismů je založena především na tradičních kultivačních postupech, které však mají schopnost detekovat pouze zhruba 1 % všech přítomných mikroorganismů. Zbývajících 99 % lze do určité míry studovat pouze za pomoci molekulárních metod. Například Hugenholtz et al. (1998) udávají, že tradičními kultivačními postupy izolujeme nejčastěji zástupce kmene Proteobacteria, Cytophagales, Actinobacteria a Firmicutes, které tvoří 90 % všech bakterií kultivovaných z půdy. Na druhé straně při klonování půdní bakteriální DNA více jak 75 % všech klonů tvoří zástupci třídy ά-proteobacteria, kmene Actinobacteria, Acidobacteria a Verrucomicrobia. Další třídy kmene Proteobacteria a kmeny Firmicutes a Planctomycetes jsou detekovány v 25–75 % studií.
Houby
Houby představují velice různorodou skupinu jak z hlediska morfologického, tak i co se týká životního cyklu. V lesních půdních ekosystémech (a i dalších fyzicky nenarušovaných půdách) předčí jejich biomasa všechny ostatní skupiny mikroorganismů a najdeme zde i zřejmě největší známý organismus zaujímající plochu 2,5 čtverečních mil, Armillaria ostoyae rostoucí v Malheur National Forest v Blue Mountains východního Oregonu. I přesto však jejich studiu v půdě je věnována podstatně menší pozornost než bakteriím a proto i naše znalosti o nich jsou značně omezené. Dosud bylo identifikováno zhruba 70 tisíc druhů hub patřících do skupin Chytridiomycota, Zygomycota, Ascomycota a Basidiomycota. Zástupci chytridií a zygomycet rychle kolonizují organické substráty přidané do půdy. Chytridie se často vyskytují v zamokřených půdách a zahrnují také parazity rostlin a živočichů. Mezi zygomycety patří i zástupci významných endomykorhizních hub. Pomalu rostoucí bazidiomycety zahrnují ektomykorrhizní houby a degradátory ligninu (Scow, 2000).

Obr. 1-6 Armillaria ostoyae
(zdroj: Wikimedia commons)
Houby v půdě rostou většinou ve formě mycelia, což jim umožňuje kolonizovat velké objemy půdy s velice heterogenně uloženými zdroji potravy. Tato jejich vlastnost má i významné důsledky pro tvorbu a udržení struktury půdy, základního předpokladu její úrodnosti. Najdeme mezi nimi mnoho patogenů rostlin, ale na druhé straně také symbiotické druhy – mykorhizní houby, jejichž význam pro výživu většiny rostlin dosud není často doceňovaný.
Zelené řasy a sinice
Také zelené řasy a výše zmíněné sinice se nachází ve většině půd, ale s výjimkou zamokřených půd zaujímají významnou roli především v prvotních fázích tvorby půdy, kde – podobně jako třeba i v pouštních habitatech – představují hlavní zdroj uhlíku a dusíku. Jak již bylo řečeno a uvádí i Tab. 1-1, najdeme v půdě i mnoho zástupců vyšších eukaryotických organismů (ale stále často mikroskopických), kteří mají v půdě svou nezastupitelnou funkci, nicméně tyto organismy už nespadají do tématu naší práce.
Až do celkem nedávné doby byly naše znalosti o složení půdních mikrobiálních společenstev založené především na tradičních kultivačních technikách, kdy byli mikrobi (především bakterie a v menší míře mikromycety) nejprve izolováni z půdy a následně kultivováni na pevných médiích. Zde však narážíme na dva fundamentální problémy: obtížnost extrakce všech mikrobů z půdního prostředí a především pak nemožnost následné kultivace většiny z nich. Podle některých odhadů lze extrahovat 33–34 % všech bakterií přítomných v půdě (Holben et al., 1988), jiné – silně optimistické – odhady hovoří sice až o 80% účinnosti extrakce (Torsvik et al., 1990a), ale i v tomto případě však kterákoliv studie používající takovou půdní bakteriální frakci nechává bez povšimnutí významnou část půdní bakteriální diverzity (v 1 g půdy může být až 1010 mikrobiálních buněk reprezentujících možná desítky tisíc různých druhů; Torsvik a Ovreas, 2002). Ještě významnější problém je již výše zmiňovaná nekultivovatelnost většiny mikroorganismů přítomných v přirozených prostředích, tedy i v půdě (Torsvik et al., 1990a; Torsvik a Ovreas 2002). Celá situace je ještě komplikovanější, vezmeme-li v úvahu také již výše zmíněnou a dosud ve většině případů zcela přehlíženou skupinu virů, k jejichž kultivaci potřebujeme hostitelskou buňku, kde – v případě bakteriofágů – opět narazíme na problém nekultivovatelnosti většiny bakteriálních hostitelů.
Z výše uvedeného je jasně patrné, že metody klasické mikrobiologie založené na izolaci a kultivaci mikroorganismů jsou nedostatečné a studium a využití obrovského potenciálu mikrobiální diverzity se neobejde bez molekulárně biologických metod využívajících nejrůznější biomarkery, tj. komponenty mikrobiálních buněk. Přehledu těchto metod využívaných ke studiu diverzity nejen půdních mikrobiálních společenstev se budeme věnovat v následujících kapitolách.
ÚEB Biol, Přírodovědecká fakulta, Masarykova univerzita |
Návrat na úvodní stránku webu, přístupnost |
| Servisní středisko pro e-learning na MU
| Fakulta informatiky Masarykovy univerzity, 2014
Centrum interaktivních a multimediálních studijních opor pro inovaci výuky a efektivní učení | CZ.1.07/2.2.00/28.0041
