Pedagogická fakulta Masarykovy univerzity Masarykovy univerzity
Martin Culek, Eduard Hofman, Boris Rychnovský, Hana Svobodová, Darina Mísařová
Biogeografické členění
Vývoj kontinentu
Austrálie byla součástí prakontinentu Pangea, který jako celek existoval beze změn od doby před 750 miliony let do doby před 150 miliony let. Indie se oddělila od Antarktidy a Austrálie před 130 miliony let (během křídy), Antarktida od Austrálie před 53 miliony let (během paleocénu) a Austrálie od Nové Guineje v raném eocénu před 45 miliony let (podrobněji viz předchozí text).
Pro další vývoj a vztahy je nezbytné si uvědomit některé skutečnosti: Z původně celistvého prakontinentu Pangea se osamostatňují jednotlivé světadíly. V severní části se rozpadá Laurasie na Severní Ameriku a Eurasii. V jižní Gondwaně přetrvává spojení Austrálie, Antarktidy a Jižní Ameriky. Ta nekomunikuje se severně položeným kontinentem – Severní Amerikou. Indie a Madagaskar jsou již samostatné (viz B). V následujícím kontinentálním rozestupovém driftu se přerušují spojovací mosty mezi Austrálií a Antarktidou a Antarktidou a Jižní Amerikou. Ta se naopak spojuje s Amerikou severní přes kontinentální most Střední Ameriky. Indie se posouvá zpět k Asii, aby se s ní nakonec spojila (viz C). Teorii kontinentálního driftu dokladují některé nálezy fosílií. Nejprůkaznější je rozpad celistvého areálu kapraďosemenné rostliny Glossopteris. Ze současných rostlin podporuje teorii kontinentálního driftu i rozšíření dalších druhů a rodů, např. pabuku Notophagus sp. v Australské a Neotropické oblasti. Tato dřevina rostla v třetihorách i v Antarktidě.
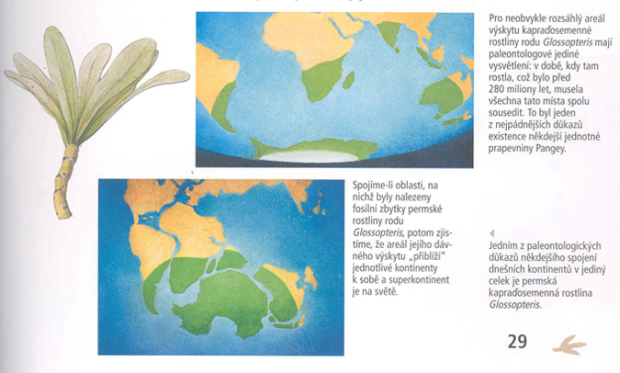
Obr. 01: Doklady příbuznosti australské bioty k biotám jiných kontinentů
Z výše uvedeného pohledu vykazuje Austrálie vysokou míru podobnosti bioty k biotám jihoamerického, antarktického a afrického kontinentu. Nezanedbatelná je i podobnost bioty s asijskou.
V rámci geomorfologického vývoje kontinentu byly prvohorní mořské usazeniny již v průběhu následujících období na rozdílných místech kontinentu vyzdvihovány a vrásněny do podoby současných pohoří. V druhohorách převážily močálové usazeniny a nánosy v deltách řek. Ze třetihor pochází uhelné močálové sedimenty, další mořské usazeniny a vyvřeliny mnohých mocných sopečných erupcí. Vrásnící pochody dotvořily povrch Austrálie do současné podoby.
Biogeografická rajonizace Austrálie
Veškerá biosféra je rozdělena na tři dílčí celky – tzv. biocykly. V každém existují rozdílné podmínky, které podmiňují způsoby života organismů.
Biocyklus:- mořský
- limnický (b. sladkých pevninských vod)
- suchozemský (b. pevninský)
Základním prostředím obou hydrosférických cyklů je voda. Do mořského biocyklu zahrnujeme vody oceánů spolu s navazujícími typy moří. Znakem je výrazná salinita – průměrná salinita oceánů činí 35 ‰. Ta limituje iontové hospodaření z pohledu izotonie nebo hypotonie tělních roztoků. Dalšími významnými faktory jsou tlak a světelné záření. Mořský biocyklus se vyznačuje výraznou kontinuitou. V mořském bicyklu rozlišujeme tři biochory: litorál a abysál s vazbou na mořské dno s hraniční hloubkou cca 200 m a pelagiál, v němž nacházíme vznášivé a plovoucí organismy bez vazby na dno. I když je rozloha limnického biocyklu svou rozlohou nesrovnatelně menší než oba ostatní, výrazná faktorová různorodost zvyšuje výslednici podmínek. Podle základního faktoru – jednosměrného pohybu vody, rozlišují mnozí autoři nejčastěji dva biochory: stojatých a tekoucích vod.
Jak napovídá název, suchozemský biocyklus zaujímá prostory mimo vodní prostředí – zahrnuje jak konkrétní místa na pevné zemi a v ní, tak i navazující vzdušné prostředí (atmosféru) až do rozsahu tolerovaném organismy. Lze ho označit i jako atmosférický. Zahrnuje rozmanité ekosystémy kontinentů, mnohdy od sebe izolovaných mořským bicyklem. Vzhledem k bohatosti životních podmínek nabízí nepřeberné množství ekologických nik.
Na základě předchozího hrubého členění jsou šelfová moře obklopující australský kontinent řazena do tropické a antiboreální oblasti. Tropická indicko-západopacifická podoblast zahrnuje většinu šelfu na východním, západním a severním pobřeží australského kontinentu. Zbývající, jižní šelf australského kontinentu označují zoogeografové jako australskou podoblast.

SV pobřeží Austrálie lemuje Velká útesová bariéra (někdy i Velký bariérový útes), nejdelší korálový útes více jak 2 000 kilometrů dlouhý a 2 – 150 kilometrů široký (Obr. 3). Podrobnější informace v boxu Bio 1. Korálové útesy mnohem menšího rozsahu leží i jihozápadně od australského kontinentu.

Obr. 03: Velký korálový útes lemuje SV pobřeží Austrálie (Kol., 2007)
Biogeografické členění pelagiálu (vodní prostředí, v němž nacházíme vznášivé a plovoucí organismy bez vazby na dno) odráží umístění kontinentů v zeměpisných pásmech. Australský kontinent včetně Tasmánie leží v tropické oblasti příslušné indopacifické podoblasti. Toto rozčlenění nezahrnuje pelagiál okolo většiny území Nového Zélandu.
Mořský abysál (prostředí mořského dna hlubší než 200 m) má v návaznosti na australský kontinent dvě části: první JV a druhá JZ a Z od kontinentu. Zoogeograficky je kategorizován do australské podoblasti antarktické oblasti.
Pevninský a limnický biocyklus vymezují obdobnou strukturu zoogeografických oblastí. Australský kontinent je centrem australské oblasti. Náleží sem i mnohé tichomořské ostrovy, ale nepatří sem mnohé blízké indonéské ostrovy (podrobněji o hranici Wallacea viz dále). Celá australská oblast obsahovala čtyři podoblasti:
- australskou kontinentální
- novozélandskou
- polynéskou
- havajskou
V dalším vývoji byla australská oblast v širším pojetí rozdělena na vlastní australskou oblast s australskou vnitrozemskou a novozélandskou podoblastí a nově koncipovanou oceánickou oblastí zahrnující početné tichomořské ostrovy včetně Havaje.
Poněkud odlišně kategorizuje oblasti fytogeografie. Rozlišuje šest floristických oblastí včetně oblasti australské.
Blízké i vzdálenější ostrovní části zoogeografické australské oblasti jsou samostatnými fytogeografickými podoblastmi paleotropu. Tropická, rozlohou menší Novoguinejská podoblast se vyznačuje tropickými lesy. Do paleotropických prvků pronikají mnohé australské elementy (blahovičníky). Výrazný je stupeň horského endemismu. Slabé vztahy k Jižní Americe jsou podmíněny existencí jižního kontinentu (Gondwany) a souvisí s kontinentálním driftem. Málo rozsáhlé (havajská, polynéská, melanésko-mikronéská, novokaledonská a novozélandská) podoblasti přes vzájemné vztahy a vztahy k mateřské indomalajské oblasti nesou vyšší stupeň endemismu.
Klima a vegetace australské oblasti
Australská oblast leží v tropickém a mírném pásu. Jih území je ovlivňován studeným Západoaustralským proudem z jižních oblastí Indického oceánu. Klimatické podmínky určují charakter přirozené vegetace.

V přímořských oblastech jsou výrazným klimatogenním činitelem východní větry vanoucí v teplé části roku z oceánu. Přinášejí srážky, které přes horská pásma poledníkového směru neproniknou do vnitrozemí (např. Blue Mountain). Obdobně v jihozápadní Austrálii se flóra vlivem bohatých srážek výrazně diverzifikovala. Klima je vhodné i k vývoji vlhkých blahovičníkových lesů. S postupným srážkovým úbytkem přechází k formám lesů tolerujících sušší klima až do chudých rostlinných společenstev suchých travin (Obr. 8).
Charakteristickým rysem klimatu jsou lijáky, působící záplavy se zdvihem hladiny o několik metrů. Výsledkem jsou dočasně zaplavovaná území a v sušším vnitrozemí i periodické řeky, vyschlé nejčastěji v chladném období (Obr. 9, 10). V tomto chladném období (květen až říjen) již vanou větry od pevniny (podrobněji v kap. Klima).
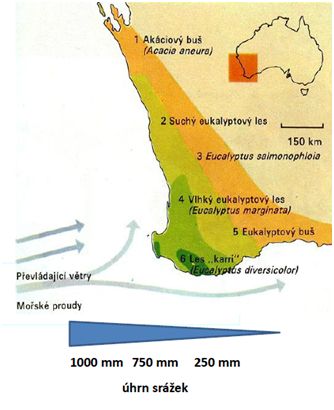
Obr. 08: Vliv větrů a srážek na různorodost lesních formací (uprav. dle Kol. 2005)

Obr. 09: Rozlehlý periodický tok („creek“)

Obr. 10: Menší „creek“ s výrazným erozivním vlivem Palmer River

Obr. 11: Mělké pánve s dočasnými zasolenými jezery. Podmínky vyhovují halofilní květeně.

Obr. 12: Zatímco halyfyty prospívají, běžné rostliny hynou

Obr. 13: Řeka Murray slouží i jako dopravní tepna

Obr. 14: Zátopová niva dolního toku řeky Murray – změna vodního režimu výrazně negativně ovlivňuje ekosystémy, které nezaplavuje a ty zůstávají bez podrostu
Vegetace australské oblasti
Flóra Austrálie je díky brzkému oddělení od ostatní pevniny a tím dlouhodobému samostatnému vývoji zcela specifická. Odhadem až 90 % rostlin je endemitních (včetně vyšších taxonů – rodů a čeledí). Druhový rostlinný endemismus je odhadován na 75 %. Rostlinstvo východní a západní Austrálie má zcela odlišný charakter – pouze 10 % druhů je společných. Logicky nacházíme fytogeografické vazby k oblastem paleotropické (východoafrické a indomalajské podoblasti) a kapské, neotropické a okrajově i holarktické, odkud pronikly středozemské elementy přes indomalajskou oblast až do australské asi na přelomu třetihor a čtvrtohor.
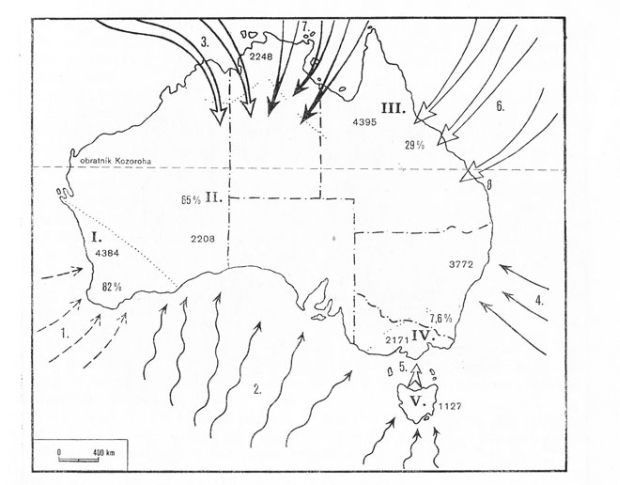
Obr. 15: Podoblasti australské vegetační oblasti a směry migrace rostlin (Brinke, 1983)
- jihozápadní – soustřeďuje asi 4 400 druhů ve vlhkých blahovičníkových a sušších kapinicových lesích.
- střední (Eremea) – suchá oblasti západní a střední Austrálie. Na největší rozlohu připadá nižší počet druhů (asi 2 200). Kromě vlhkých blahovičníkových a sušších kapicových lesů doznívajících ze západní strany, jsou pro suchém vnitrozemí charakteristické savany, stepní formace až polopouště a pouště. Na nich rostou společenstva tvrdých ostnitých trav – tzv. spinifexové formace.
- severovýchodní – s tropickým deštným lesem v tropické části a navazujícími monzunovými lesy s ubývajícími teplotami směrem na jih. S úbytkem srážek přechází v sušší travnaté formace. Hranice s jižnější podoblastí je totožná s hranicí mezi státy Nový Jižní Wales a Victoria. V podoblasti mají výrazné zastoupení indomalajské prvky – někteří autoři přiřazují úzký pobřežní pruh S Austrálie do Indomalajské podoblasti v rámci Palearktické oblasti. Vysokým počtem druhů je podoblast rovna jihozápadní.
- jihovýchodní – zahrnuje jednak horskou, jednak pobřežní část jihovýchodní Austrálie. Od pobřeží do hor přechází suché blahovičníkové lesy do podoby subalpínské až alpínské vegetace Oproti ostatním je zřejmý úbytek rostlinných druhů (asi 2 200).
- tasmánská – navazuje a tím se velmi podobá předchozí. Charakterizuje ji další úbytek druhů (asi 1 200 druhů).

Obr. 16: Blahovičníky horské (E. regnans) patří k nejvyšším stromům planety (NP Dandenong)

Obr. 17: Akácie rostou nejčastěji v polosuchém vnitrozemí
Z předchozího textu vyplývá vyšší náročnost blahovičníků na množství srážek. Z obr. 18 je patrná nejvyšší frekvence jejich rozšíření ve vlhčích podoblastech severní, východní a jihozápadní Austrálie. Při zahrnutí suchomilnějších akácií do hodnocení je četnost výskytu obou rodů nejvyšší v celém východním pásu Austrálie a v jihozápadní návětrné části (Obr. 19) nejpočetnějších australských stromových forem (blahovičníků a akácií). Na ostatních biotopech s nižšími srážkami se díky úbytku blahovičníků tyto druhy vyskytují sporadičtěji.
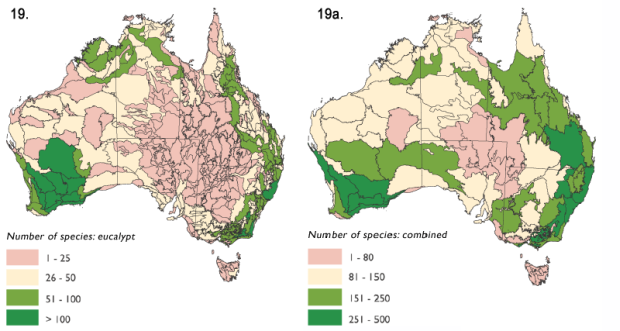
Obr. 18 / 19: Biodiverzita blahovičníků v rámci subregionů Austrálie / Biodiverzita blahovičníků a akácií v rámci regionů Austrálie (www.gov.au)
Z nahosemenných rostlin zde nacházíme blahočety (Araucaria sp.), cykasy (Macrozamia sp.), damaroně (Agathis sp.) aj.

Obr. 20: Typickými australskými dřevinami jsou také blahočety

Obr. 21: Cykasy Macrozamia mcdonellii rostou v centrální Austráli
V tropické oblasti, tj. na severu a SV Austrálie a Nové Guinei rostou, díky vlhkému a teplému klimatu, tropické lesy (Obr. 22, 23). Na menším území s průměrnými ročními srážkami nad 2000 mm se zde vyskytují zbytky tropického deštného pralesa. Výrazné zastoupení v nich mají banánovníky (Musa sp.), blahovičníky (Eucalyptus sp.), liánová palma rotang (Calamus sp.) a další, ale i stromové i epifytické kapradiny. Podnebí Nové Guiney se oteplilo v relativně nedávné minulosti. Z té doby pocházejí některé relikty jako blahočet Cunninghamův (Araucaria cunninghami), fylokladus Phyllocladus hypophylla a rozpylec lékařský (dříve pepřovník čilský) Drimys winteri. Část novoguinejské flory (i fauny) se shoduje s flórou (i faunou) severoaustralského Queenslandu v důsledku častého spojení souše při poklesu hladiny šelfového moře. Ostrov je výrazně hornatý, existující alpínské pásmo na jednotlivých horských hřebenech je mozaikovité.

Obr. 22: I v australském tropickém lese je většina metabol. pochodů soustředěna do korunového patra

Obr. 23: Nápadným stromem je kvetoucí brachychiton javorolistý(Brachychiton acerifolium)
Deštný tropický les SV Austrálie v závislosti na množství srážek přechází na blahovičníkové nížinné lesy a horské vlhké lesy (Obr. 24). Spolu s vlivy klimatu se mění v jižním směru na lesy mírného pásma.

Obr. 24: Blahovičníkové lesy v Blue Mountains, foto: Čech, T., 2010
Mangrove, porosty vždyzelených dřevin na kontaktu moře a souše, rostou v tropickém a subtropickém pásu v přílivové zóně (zóně vymezené přílivem a odlivem – Obr. 25). Nepříliš četní zástupci (asi 20 druhů dřevin jako jsou kolíkovník Avicennia sp., kořenovník Rhizophora sp., kuželovník Sonneratia sp.) mají schopnost přijímat vodu ze zasoleného prostředí – dokáží zvýšit osmotický tlak v buňkách nad tlak okolní mořské vody.

Obr. 25: Zonace mangrovových porostů (upraveno podle Jeník, 1996)

Obr. 26: Významný mangrovový druh – kořenovník (Rhizophora sp.)
Periodicky kolísající hladině vody se přizpůsobují rostliny jednak chůdovitými (Obr. 26), jednak dýchacími kořeny. Ty současně pomáhají pevnějšímu ukotvení jedince v bahnitém substrátu a naopak přispívají k ukládání sedimentů. Na mangrovové porosty navazuje zasolený pás řídké halofytní vegetace až k úrovni skočného přílivu (Obr. 27). Mangrovové porosty chrání pevninu před poškozujícím vlnobitím (včetně tsunami) a podporují rozšiřování pevniny. Kromě běžných živočichů se specifickým životním podmínkám přizpůsobily i mnohé taxony zajímavých živočichů (noroví krabi, hlaváčovité ryby – lezci /Obr. 28/ a další). Mangrovové porosty lemují nepravidelně severní, východní a jihovýchodní pobřeží Austrálie.

Obr. 27: Celkový pohled na mangrovové porosty (Cairns)

Obr. 28: Lezec bahenní (Periophthalmus argentilineatus) – výrazný obyvatel mangrovů
V lesích mírného pásu převažují pabuky r. Nothofagus. Dalším významným druhem vlhkých oblastí JV Austrálie jsou blahovičníky horské. Ve vlhkém podrostu rostou stromové kapradiny (r. Dicksonia sp. nebo Cyathea sp. – Obr. 16). Relativně bohaté srážkové poměry JV Austrálie zde umožňují růst rostlinstva v téměř džunglových podmínkách. Lesy podobné skladby nacházíme na Tasmánii i Novém Zélandu. V západní části kontinentu se arboreál nachází omezeně pouze na JZ (viz Obr. 8). Zbytky původních lesů jsou chráněny v národních parcích (podrobněji viz Ochrana přírody).
Horské lesy typicky rostou ve vlhkých podmínkách Snowy Mountains (Sněžné hory). Ty jsou součástí Velkého předělového pohoří. Vyhlášený národní park (Kosciuszko National Park) pokrývá největší australskou oblast alpinského charakteru. Kromě něj byly vyhlášeny další národní parky Australských Alp v propojeném systému (Brindabella NP, Namadgi NP, Snowy River NP, Alpine NP a Mt. Buffalo NP – podrobněji viz Ochrana přírody). Ve Snowy Mountains se rovněž nachází nejchladnější místo australského kontinentu. Ve vyšších polohách (jedenáct vrcholků nad 2 000 metrů nad mořem s nejvyšší horou pohoří i celého kontinentu Mt. Kosciusko – 2 228 metrů nad mořem) trvá dlouhá sněžná a mrazivá zima. Blahovičníkové lesy (Obr. 29, 30, 31) s typickým stromem blahovičníkem sněžným (Eucalyptus pauciflora subsp. niphophila) jsou charakteristické pro subalpinské pásmo v rozmezí 1 500 až 2 000 m n. m. Ve vyšších nadmořských výškách přechází v typické alpinské pásmo (Obr. 32). Rostlinná společenstva tady tvoří mozaikovitý porost krátkostébelných travin, nízkých bylin, keříků a vřesovišt (viz Box Bio 3).

Obr. 29: Typické horské lesy s blahovičníkem sněžným v Snowy Mountains

Obr. 30: Vnitřek prosvětleného lesního porostu blahovičníku sněžného (Eucal. paucifl. niphophila) s keřovým podrostem

Obr. 31: Blahovičník sněžný (Eucalyptus pauciflora niphophila) na Mount Hotham (1786 m n.m.)

Obr. 32: Kvetoucí horská krajina alpínského pásma pod Mount Kosciuszko (s jezerem Cootapatamba)
Až 70 % území centrální a západní Austrálie zahrnují pouště, polopouště a suché stepi. To zapříčiňuje horská východní bariéra, zadržující vlhké JV pasáty a na opačné straně ochlazené západní větry bez patřičné vlhkosti. I když pouze část suchého vnitrozemí je skutečnou pouští, rozlišujeme pět typů (Obr. 33, 34):
- pískové p.
- kamenité p.
- hornaté p.
- tabulové p.
- hlinité p.

Obr. 33: Písková poušť centrální Austrálie

Obr. 34: Kamenitá poušť centrální Austrálie (Henbury)
Nejsou zcela bezsrážkové – minimální srážky okolo 100 mm ročně umožňují kromě travin na příhodných místech v dolinách růst i stromů (přesličníky a akácie). Polopouštní podmínky snášejí přesličníky (Allocasuarina sp. – Obr. 35). Na suché podmínky jsou mnozí živočichové (někteří ještěři aj.) dostatečně adaptováni nokturnální aktivitou, minimalizací potřebného množství vody i jejích ztrát.

Obr. 35: Přesličník Decaisneův (Allocasuarina decaisneana) roste v centrální Austrálii, jeho příbuzní i v Malayasijské oblasti
V suchých podmínkách centrální Austrálie tvoří hlavní rostlinné formace tvrdolisté křoviny. Obecně se označují jako scrub (brigalow-scrub). Tam, kde převažují několik metrů vysoké porosty blahovičníků, užívá se název mallee, křoviny s převahou akácií jsou mulga. Na zasolenějších půdách je charakteristickým útvarem tzv. „lebedová step“ se zastoupením merlílkovitých rostlin (Atriplex hymenelytra – nízký halofytní a suchomilný keř, merlík trpasličí – Chenopodium pumilio).

Obr. 36: Porost tvrdolistých křovin – brigalow-scrub

Obr. 37: Formace tvrdolistých dřevin s převahou keřovitých blahovičníků je nazývána „mallee“

Obr. 38: Akácie jsou základním prvkem křovitých formací „mulga“

Obr. 39: Ze suchomilných pestře kvetoucích rostlin zaujme nádhernice úhledná (Clianthus formosus), endemit centrální Austrálie

Obr. 40: Ptilotus obovatus roste v aridních oblastech v kamenitých půdách
Australská zoogeografická oblast (Australasie)
Australská zoogeografická oblast zahrnuje celý australský kontinent s přilehlými ostrovy Celebesem (Sulawesi) a Novou Guineou na severu a jižně položenou Tasmánií, které výrazněji vystupují ze šelfového území a mnohé další méně rozlehlé ostrovy. Řazení hornatých ostrovů k Austrálii spočívá v tektonických souvislostech. Zatopení rozlehlého kontinentálního šelfu a oddělení ostrovů proběhlo v průběhu posledních 10 000 let. V oblasti Sundského moře (navazuje na Novou Guineu) sousedí australská s indomalajskou oblastí. Na jednotlivých ostrovech se prolínají prvky indomalajské oblasti s australasijskou v různé míře. Přesné stanovení hranice není téměř možné. Bylo zde definováno několik hraničních linií – nejčastěji je upřednostňována hranice ve smyslu tzv. Wallaceovy linie (Obr. 41). Podrobnější vymezení hraniční oblasti naleznete v Boxu Bio 4 (Obr. 42).
Dalším ostrovem, vývojově odděleným hlubokým oceánem, řazeným do australské oblasti je Nový Zéland. Celková rozloha australské zoogeografické oblasti: 9 mil. km2.
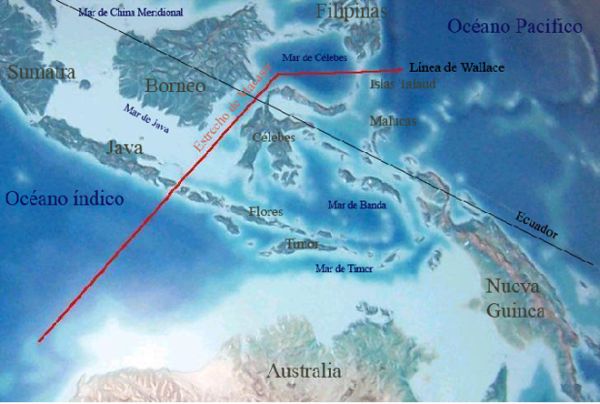
Obr. 41: Nejčastější interpretace hranice indomalajské a australasijské biogeografické oblasti (zdroj)
Kromě australské flóry a fauny se na Novou Guineu v různých dobách rozšířilo mnoho zástupců z JV Asie, zvláště ptáků. Z toho důvodu je rozmanitost ptačích zástupců na rozlohově výrazně menší Nové Guinei stejná jako ve výrazně rozlehlejší celé Austrálii. Přesto zde některé ekologicky významné skupiny chybí a jsou vikarizovány skupinami jinde neznámými (vikarizace datlů drobnými papoušky). Významnou adaptovanou skupinou ptáků jsou rajky s tokem založeným na vizuální komunikaci. K tomu mají polygamní samci extrémně vyvinuté a zbarvené peří. Čtyři druhy pronikly i do lesů severní Austrálie.
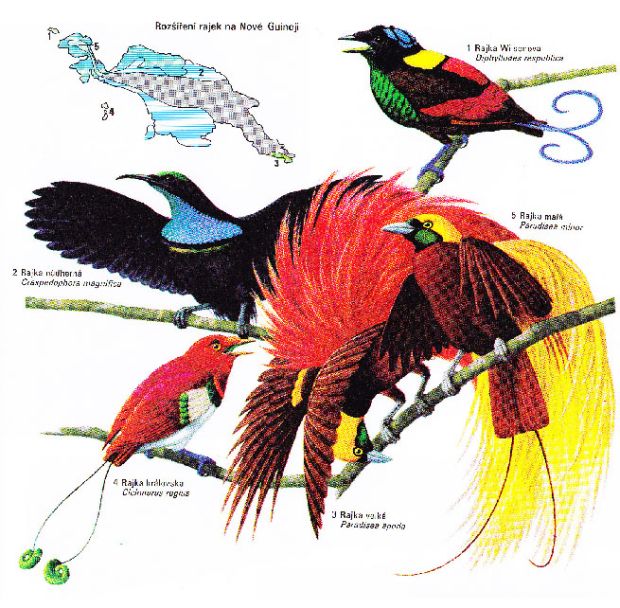
Obr. 43: Příklad barevnosti rajek a rozšíření na Nové Guinei (Kol., 2005)
Příbuzní rajek, lemčíci, žijí obdobně v lesích Nové Guineje a severní Austrálie. Jejich tok je založený na budování stavby v podobě besídky nebo loubí samcem. Tuto stavbu doplňuje tvarově a barevně nápadnými předměty (včetně odpadů).
Další vysoce zajímavou skupinou celé indonésko-australské oblasti jsou taboni – nekrmiví ptáci inkubující vejce bez sezení rodičů tepelnou energií prostředí. K tomu využívají buď infračervenou složku slunečního záření v interakci s tmavým sopečným pískem, geotermální energii sopečných oblastí nebo zbytkovou tepelnou energii rozkládajícího se rostlinného materiálu. Některé druhy žijí v deštných lesích, některé v sušším prostředí (tabon holubí).

Obr. 44: Tabon lesní (Alectura lathami) z tropických lesů severní Austrálie se výrazně synantropizuje
Fauna Austrálie včetně Nové Guiney a Tasmánie se vyznačuje výrazným druhovým zastoupením savců podtřídy vejcorodých a vačnatců z třídy živorodých. To je výsledkem dlouhodobé izolovanosti kontinentu od ostatních. Vývoj fauny se zde rozvíjel bez souvislostí s ostatními kontinenty, což se zřetelně projevilo v adaptivní radiaci vačnatých savců neovlivňované progresivnějšími placentály. Výhradně v této oblasti nacházíme zástupce vejcorodých savců: ptakopyska podivného (Ornithorhynchus anatinus), ježury australské (Tachyglossus aculeatus) a paježury. Jejich mláďata se líhnou z vajec, která ptakopysk zahřívá v hnízdě, zatímco ježury nosí ve zvláštním vaku. Ptakopysk se vyznačuje jakoby „kachním“ zobákem, veslovitým ocasem a plovacími blánami mezi prsty. Zatímco výskyt ptakopyska je pouze mozaikovitý (sladké vody východní Austrálie a Tasmánie), ježury zasídlují většinu biotopů jmenované oblasti včetně Nové Guineje.

Obr. 45: Spatřit ptakopyska ve volné přírodě není lehké

Obr. 46: Ježura australská se živí převážně termity a mravenci
Další australští autochtonní savci – vačnatci nepřekračují Wallaceovu linii. V Australské oblasti žije většina (70 %) vačnatců z celkem 170 druhů. Z nich je nejpočetnější a pravděpodobně i nejznámější čeleď klokanovitých (56 druhů podle Veselovského 1986, 53 druhů dle Wikipedie, přičemž některé druhy jsou hodnoceny jako vymřelé). Velikostně i topicky jsou tito býložravci značně rozrůzněni: největší zástupci přesahují svojí výškou člověka (klokan obrovský, k. rudý), naopak nejmenší dosahují velikosti potkana, tj. 100násobně nižší hmotnosti (Box Bio 5). Většinou jsou klokani stepními druhy, ale adaptovali se životu ve skalním terénu (k. skalní) i na stromech (k. pralesní a k. stromový). Menší druhy jsou obecně označováni jako walabi (klokan rudokrký, k. bažinný). V hustěji zalidněných a hospodářsky využívaných oblastech byli výrazně redukováni.

Obr. 47: Klokan velký (Macropus fuliginosus) patří k velkým druhům
V druhé nejpočetnější čeledi australských vačnatců (kunovcovití) jsou zastoupeny různě adaptované a vzhledově rozdílné, většinou dravé druhy. Byli srovnáváni se známými evropskými placentárními savci (kunovci, vakorejsek, vakomyš, vakotarbík, případně mravencojed). Zástupci méně početných čeledí, vakoveverky, vakoplši a vakokrti připomínají svými charakteristikami opět dříve známé zástupce Starého světa. Všežraví bandikuti tvoří přechod k býložravým kuskusům a possumům. Asi nejpopulárnějším vačnatcem je koala (Phascolarctus cinereus) – oligofágní až monofágní býložravý potravní specialista, žeroucí listy vybraných druhů blahovičníků (Box Bio 6). Hrabaví vombati svým zavalitým vzhledem evokují neohrabaná zvířata. Vykazují převážně noční aktivitu jako většina předchozích. Dalším, tentokrát negativně proslulým vačnatcem je vakovlk neboli tasmánský tygr (Thylacinus cynocephalus). Tento dravý vačnatec velikosti středního psa byl z Nové Guineje a Austrálie pravděpodobně vytlačen dingem. Přežíval na Tasmánii, kde ho ve velkém v krátkém čase vyhubil člověk. Vačnatce nacházíme i na kontinentu Jižní Ameriky (asi 70 druhů), odkud v nedávné době pronikli až do Ameriky Severní (vačice).

Obr. 52: Koala je potravním specialistou

Obr. 53: Noční norový vačnatec vombat obecný (Vombatus ursinum)
Při hodnocení cennosti australského území z pohledu bohatosti endemických savců (Obr. 40) vyniká Yorkský poloostrov na severovýchodě Austrálie. Východní pobřežní pruh a malá území Kimberley jsou oblastmi středního významu pro výskyt endemických savců. Rozsáhlé území zbytku Austrálie včetně jihozápadu nenese pro endemické savce takový význam.
Mezi menšinu placentárních savců patří dingo (pes dingo) (Canis dingo), introdukovaný původními domorodci při jejich imigraci do Austrálie (asi před 3 – 5 tisíci let – Anděra, 1997), mořští ploutvonožci, drobní hlodavci a letouni.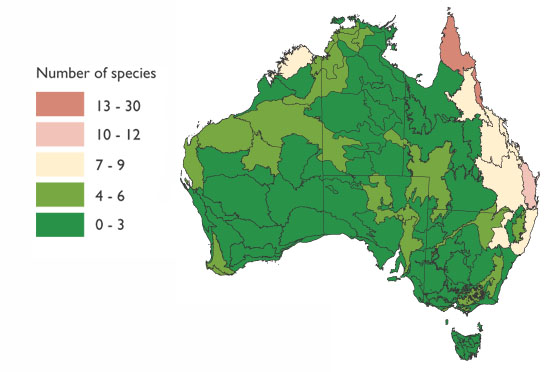

Obr. 55: Vyjící dingo
Zástupci placentárních savců jsou netopýři, kaloni a krysy (čel. Muridae). Zástupci myšovitých jsou děleni na „staré endemity“, jejichž přistěhování proběhlo před 5 – 10 miliony let a „nové endemity“, u nichž vývoj na kontinentu trvá asi jeden milion let. K nezáměrně introdukovaným jsou řazeny nejvýznamnější imigranti: myš domácí, potkan a krysa (en.wikipedia.org/Australasia). Výlučně mořské savce ve vodách obklopujících Austrálii (z sirén moroně Dugong dugong a kytovce – celkem 59 druhů) řadíme do mořského biocyklu.

Obr. 56: Den tráví kaloni šedohlaví (Pteropus poliocephalus) v klidu ve větvích

Obr. 57: Kaloni vyletují večer za potravou (sladké plody). V průběhu dne mění své odpočinkové místo v důsledku soupeření.
Australskou oblast charakterizují také endemičtí běžci: největším ptákem oblasti je emu hnědý (Dromaeus novaehollandiae), v současnosti vytlačený ze zemědělských oblastí do méně příhodných podmínek otevřené suché krajiny. Menší lesní příbuzný, kasuár přílbový (Casuarius casusarius), obývá deštné lesy severní Austrálie a Nové Guineje. Je nápadný kostěným hřebenem a zbarvením krku.

Obr. 58: V keřových až travinných biomech australských polopouští žije emu hnědý

Obr. 59: V deštných lesích Nové Guineje a severní Austrálie žije kasuár přílbový
Jižní pobřeží Austrálie a dalších ostrovů obývá jeden z nejmenších tučňáků – tučňák nejmenší (Eudyptula minor). Hnízdí v norách vyhrabaných v pobřežních píscích.

Obr. 60: Tučňák nejmenší se večer vrací ke krmení mláďat v norách

Obr. 61: Labuť černá

Obr. 62: Orel klínoocasý funguje jako mrchožrout
I když zástupci papoušků žijí i na ostatních kontinentech, pro Austrálii je typická čeleď kakaduovitých (Cacatuidae). Je na nich nápadná vztyčitelná chocholka. Někteří zástupci jsou tak četní, že hledáním potravy mohou poškozovat i zemědělskou produkci. Mezi populární klecové ptáky řadíme korelu (Nymphicus hollandicus). Dalšími původně endemickými papoušky Austrálie jsou rosely, loriové a andulka vlnkovaná (Melopsittacus undulatus) – jeden z nejčastěji chovaných ptáků. Z klecových chovů se rekrutují výskyty mimo původní oblast (včetně ČR).

Obr. 63: Početný kakadu žlutočečelatý (Cacatua galerista)

Obr. 64: Kakadu růžový (Cacatua roseicapilla) není farmáři oblíben

Obr. 65: Rosela Pennantova (Platycercus elegans) si snadno zvykla na člověka

Obr. 66: Lori mnohobarvý (Trichoglossus haematodus) se živí nektarem
Ledňáci jsou středně velcí ptáci, lovící hmyz a drobné obratlovce v řidších lesích. Ledňák obrovský se mnohdy přiživuje až loupí na příležitostné potravě lidí. Jeho volání patří k nápadným a častým hlasům australské přírody.

Obr. 67: Ledňák modrokřídlý (Dacelo leachi) je téměř tak velký jako l. obrovský
Mezi charakteristické ptáky Austrálie patří kromě jiných již zmiňovaní zástupci tabonovitých, rajkovitých a lemčíkovitých. Výrazný ocas lyrovitého tvaru se dvěma typy ocasních per vlastní lyrochvost (velechvost) nádherný (Menura novaehollandiae). Funkcí nápadných ocasních per je toková prezentace doplňovaná výraznými hlasovými projevy. Další charakteristická čeleď australských pěvců – kystráčkovití (Meliphagidae, někdy medosavkovití), opyluje kvetoucí rostliny při sběru nektaru jako jihoameričtí kolibříci. Stejnou funkci plní i strdimilové, zastoupení hlavně v africké oblasti. V ornitofauně Austrálie chybí někteří u nás běžné taxony: sýkory, ťuhýci i čeleď pěnkavovitých pěvců. Ti jsou nahrazeni zrnožravými snovači (Ploceidae). Mnozí pestře zbarvení jsou chováni v klecích – nejznámější je asi páskovník šedý (Poephila guttata), dřívějším názvem zebřička pestrá (Taeniopygia castanotis).

Obr. 68: Páskovník šedý (zebřička pestrá) – běžný zrnožravec Austrálie

Obr. 69: Nektarem a hmyzem se živí strdimil žlutý (Nectarinia jugularis)
Endemickým krokodýlem Austrálie je krokodýl australský (Crocodylus johnsoni). Známější a člověku nebezpečný je krokodýl mořský (Crocodylus porosus). Sice vyplouvá daleko do oceánu, ale žije i v lagunách se sladkou vodou. Délkou 7 m je největší recentní krokodýl.

Obr. 70: Krokodýl mořský v posledních letech díky ochraně rozšířil svůj areál
Ještěři jsou zastoupeni mnohými gekony (z endemických čeledí Diplodactylus, Oedura, Nephrurus) a dalšími s širším rozšířením. Endemická je i šupinonožka černohlavá (Pygopus nigriceps) – hadovitý gekon se zvláštními zadními končetinami v podobě šupiny. Výrazně je zastoupena čeleď scinkovitých (rody Egernia, Ctenotus, Tiliqua a další).

Obr. 71: Egernie (Egernia major) obývá lesní biotopy i antropogenní biotopy

Obr. 72: Živorodý scink uťatý (Trachydosaurus rugosus) má primitivní žloutkovou placentu
Největší agamou v Austrálii je agama vodní (Physignathus lesueri). Je vázaná na biotopy okolo vod, dobře plave. Pro sušší oblasti severního území je typická agama límcová (Chlamydosaurus kingi) se širokým kožním záhybem pro zastrašování protivníků. Menší agamou, typickou pro pouštní biotopy je moloch ostnitý (Moloch horidus).

Obr. 73: Agama vodní je nápadná velikostí

Obr. 74: Moloch ostnitý patří k menším ještěrům (velikosti do 20 cm)
Centrum rozšíření varanů se nachází v širší hraniční oblasti, jenom na australském kontinentu žije 26 z asi 50 druhů (en.wikipedia.org/Fauna of Australia). Nejběžnějším varanem je varan Gouldův (Varanus gouldii).

Obr. 75: Často jsou jedinci varana Gouldova usmrceni provozem na silnicích
Hadi mají zastoupení v čeledích slepákovití (Typhlopidae), hroznýšovitých (Boidae), převážně jedovatých korálovcovitých (Elapidae) a užovkovitých (Colubridae), zastoupených druhy nejedovatými i jedovatými (bojgy). Jako nejjedovatější had je hodnocen taipan (Oxyuranus scutellatus). I sladkovodní želvy jsou převážně endemické.
Obojživelníci jsou zastoupeni žábami (rosnice a hvízdalky), ocasatí a červoři chybí.

Obr. 76: Stromová rosnice siná (Litoria caerulea)
se částečně synantropizovala
Limnický biocyklus je nápadně chudý – mezi primárně sladkovodní ryby řadíme jediný druh – bahníka australského (Neoceratodus forsteri). Jeho výskyt je plošně velmi omezený, i když jeho druhohorní předkové měli kosmopolitní rozšíření. V současnosti žije ve sladkých vodách asi 170 druhů. Z nich 70 % má příbuzenské vztahy k mořským indo-pacifickým předkům, kteří se sekundárně adaptovali na vody sladké. Nejznámější je baramundi (Lates calcalifer). Mnohé sladkovodní nepůvodní druhy byly do vod introdukovány (pstruzi, losos atlantský, jeho tichomořští příbuzní r. Oncorhynchus, středoevropský kapr obecný, živorodka komáří). Z nich jako problémoví jsou hodnoceni kapr a živorodka komáří. Druhová rozmanitost ryb je vysoká – mořské vody okolo Austrálie obývá odhadem 1 500 druhů.
Zastoupení všech bezobratlých na australském kontinentu se odhaduje asi na 190 000 druhů. Zatím je známa asi polovina. Setkáme se s nimi ve většině ekosystémů. V taxonech hmyzu a měkkýšů je 90 % hodnoceno jako endemických. Z pavouků endemický sklípkan (Atrax robustus) žije pouze v oblasti Velkého Sydney. Nejpočetnějším taxonem je hmyz (75 % známých australských živočichů). Nediverzifikovanějšími řády jsou brouci a motýli (28 200 resp. 20 816 druhů).

Obr. 77: Mravenec tkalcovský (Oecophylla smaragdina)
sešívá listy pro hnízdo ve větvích stromů
Mnohé ostrovy australské oblasti mají faunu odlišnou od fauny Austrálie. Celebes (Sulawesi) je krajním ostrovem australské strany Walaceovy linie. Vačnatci jsou zde zastoupeni kuskusovitými druhy (Phalangeridae). Míra zastoupení placentálních savců je výrazně vyšší než ve zbytku australské oblasti – poloopice jsou zde zastoupeny unikátním nártounem okatým (Tarsius spectrum), vyšší opice makakem tmavým (Macaca nigra). Endemickými sudokopytníky jsou 1 m vysoký buvol anoa (Bubalus depressicornis) a babirusa (Babyrousa babyrussa) z příbuzenství prasat.
Na Nové Guineji jsou zastoupeni vejcorodí savci (ježura a paježury), výrazněji než v předchozím případě i vačnatci (kromě kuskusů několik druhů bandikutů a klokanů). Placentálové jsou omezeni na hlodavce, letouny a ze šelem psa pralesního (Canis hallstromi), možné subspecie dinga. Hlavní centrum rozšíření zde mají rajky, lemčíci a holubi-korunáči. Endemitním plazem je krokodýl novoguinejský (Crocodylus novaeguineae), zatímco areál taipana (Oxyurans scutellus) zahrnuje Novou Guineu i Austrálii. Mnohé rosničkovité (Hylidae) a parosničkovité (Microhylidae) žáby jsou endemity (Opatrný, 2001).
Fauna Tasmánie je velmi podobná australské. Žije zde ďábel medvědovitý (Sarcophilus harrisi), zatímco vakovlk psohlavý (Thylacinus cynocephalus) byl pravděpodobně vyhuben, stejně jako místní forma emu.
Ve fauně Nového Zélandu s přilehlými menšími ostrovy téměř chybí plazi a savci (pouze několik druhů scinků a gekonů, hatérie novozélandská (Sphenodon punctatus), 2 druhy netopýrů – (Mystacina tuberculata) endemický, krysa maorská (Rattus exulans) a endemický lachtan (Neophoca hookeri). Významnou složkou jsou nelétaví ptáci – ze 3 endemických čeledí jsou nejznámější kiviové. Domorodí Maorové vyhubili odhadem v 10. až 14. století až 3 m velké nelétavé ptáky moa (Dinornithis moa). Mezi endemické bezobratlé patří mimo jiné i čeleď motýlů s kusadly Mnesarchidae. Mezi rostlinami řadíme většinu k endemickým. Na rozdíl od vlastního australského kontinentu se zde nevyskytují blahočety Araucaria sp.
I když je fauna pacifických ostrovů (mj. Havajské souostroví, Fidži, Tongo) silně endemická, od australské se výrazně liší hlavně nepřítomností ptakořitných a vačnatých savců.
Z hlediska druhové bohatosti je nejvýznamnější fauna Nové Guiney (ze 175 druhů savců řadíme 128 mezi endemity ostrova) a severní Austrálie. Naproti tomu je fauna Nového Zélandu a tichomořských ostrovů nápadně chudší. Proto zde přežily bez rizika konkurence některé taxony, které jinde vyhynuly (např. hatérie). Blízké australské ostrovy jsou floristicky i faunisticky vlastní oblasti mnohem bližší. Mezi nimi je významný ostrov Fraser i Ostrov lorda Hovea pro zachovalost přírodních podmínek a mnohé rostlinné i živočišné endemity (viz Ochrana přírody).
Invazní rostliny a živočichové
Vážný hospodářský problém způsobila opuncie (Opuntia robusta), introdukovaná z jižní Ameriky. Svým rychlým šířením (v maximu až 200 000 km2 hlavně ve východní Austrálii v oblasti Brisbane) i na úrodných půdách se stala hrozbou pro zrninové zemědělství. Běžné způsoby likvidace byly málo účinné – až využití hmyzího fytofága (housenky Castoblastis sp.) zastavilo další šíření.
Všechna důležitá hospodářská zvířata byla do Austrálie přivezena. Mnohá z nich nenašla v nových podmínkách patřičné regulační mechanismy a jejich nekontrolovatelné rozmnožování vyústilo do škodlivého početního nárůstu. Zářným příkladem nebezpečí neřízené introdukce byl dovoz a vypuštění králíka divokého (Oryctolagus cuniculus).

Obr. 78: Králík divoký se stal pohromou travinných oblastí Austrálie
Postupnou kolonizaci od introdukce v r. 1859 zachycuje Obr. 79. Začátkem 50. let byla početnost populace králíka vyčíslena na 4 miliardy jedinců. Šíření králíka se podařilo omezit až uměle naočkovaným nákazovým onemocněním – myxomatózou. I malá přežívající část imunní populace byla schopna zakládat ohniska opětovného strmého populačního růstu (Brinke, 1983).

Obr. 79: Šíření králíka v Austrálii a rozšíření psa dinga (Foltýnová, 2003)
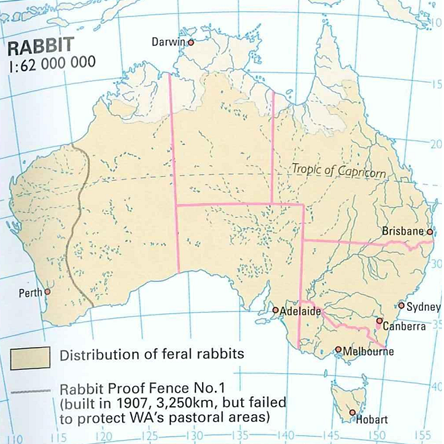
Obr. 80: Výskyt králíka divokého na australském kontinentu (Kol., 2007)
Dalšími příklady škodlivé záměrné introdukce byli timorští vodní buvoli (Bubalus bubalis). Z pouhých několika kusů v roce 1807 se současná populační početnost odhaduje na 250 000 jedinců. Nižších počtů dosahuje zdivočelý domácí kůň (165 000). Z toho pohledu početní stavy velbloudů, původně introdukovaných ke karavanní dopravě v suchých vnitrozemských polopouštích a pouštích, nedosahují zdaleka významu předchozích.

Obr. 81: Významnými býložravými konkurenty původní fauny Austrálie se stali koně

Obr. 82: Býložravými konkurenty původní fauny Austrálie se stali i velbloudi jednohrbí
Dalšími příklady nevhodné introdukce s následným škodlivým přemnožením jsou lišky (Obr. 83) a vrabci. Méně známé introdukce se týkají dalších býložravců (jeleni), případně všežravců (prase divoké).

Obr. 83: Liška obecná (Vulpes vulpes) (introdukovaná 1855 jako lovné zvíře) zasídlila přibližně stejné území jako králík (Kol., 2007)
Posledním negativním případem je introdukce ropuchy obrovské (Bufo marinus). S ropuchou obrovskou jsme se setkali v noci vysoko v horách na cestě deštným lesem. Pochází ze střední a jižní Ameriky. Poznáme ji podle výrazných parotidálních žláz za očima (viz obr. 84). Byla do Austrálie v roce 1935 přivezena k likvidaci škůdců (listožravých brouků) na plantážích cukrové třtiny. Počáteční předpoklady se nenaplnily, oba taxony nevstoupily do očekávaného potravního řetězce.

Obr. 84: Introdukovaná ropucha obrovská
Prospívají jak brouci, tak i ropuchy, které se následně se rozšířily i do původních biotopů tropického lesa (Obr. 86). Zde se přeorientovaly na potravu v podobě drobných původních živočichů. Pro většinu potenciálních australských predátorů je pro jedové výměšky nepoživatelná, dokonce jedovatá a výsledkem jsou jejich otravy. I přes neustálé hubení není prakticky možnost jejího výrazného omezení.

Obr. 85: Hospodářské porosty cukrové třtiny
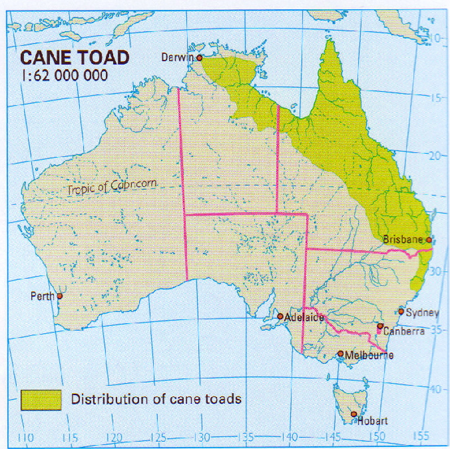
Obr. 86: Současné rozšíření zavlečené ropuchy obrovské v Austrálii (Kol., 2007)
doc. RNDr. Boris Rychnovský, CSc. a kolektiv |
KGeog, Pedagogická fakulta, Masarykova univerzita |
Návrat na úvodní stránku webu, přístupnost |
| Servisní středisko pro e-learning na MU
| Fakulta informatiky Masarykovy univerzity, 2013
Centrum interaktivních a multimediálních studijních opor pro inovaci výuky a efektivní učení | CZ.1.07/2.2.00/28.0041
